Giant clams have long fascinated adventurers and naturalists. These large shallow-water molluscs certainly are among the most colourful, conspicuous and emblematic species of the Indo-Pacific coral reefs. They have been exploited for thousands of years for their flesh and shell. Giant clam conservation is also an increasingly concerning issue because of the vulnerability of giant clams to overharvesting. Surprisingly, up to recent years, giant clams have remained incompletely known and described, and their evolutionary history was poorly understood.
Image: C. Fauvelot (IRD) is doing a biopsy on a giant clam in Juan de Nova. Photo credit: T.B. Hoareau / TAAF-Iles Eparses research consortium.
———————— FROM THE COVER: read the article on which this post is based … Fauvelot, C, Zuccon, D, Borsa, P, et al. 2020. Phylogeographical patterns and a cryptic species provide new insights into Western Indian Ocean giant clams phylogenetic relationships and colonization history. J Biogeogr. 47:1086– 1105. https://doi.org/10.1111/jbi.13797 ————————
As Tridacna giant clams exclusively occur in coral reefs of the Indo-West Pacific (IWP), we believed that studying the mode and tempo of their speciation would provide us with clues on the evolutionary history of modern coral reef communities in the IWP. We addressed this objective by combining molecular phylogenies with the geographic distribution of Tridacna lineages across the IWP. With well-dated, albeit rare fossil records, we had a model of choice to link phylogenetic patterns to past geological events.
When we started our phylogeographic research in the late 2000s, little was known from the Indian Ocean although a robust phylogeography of Coral-Triangle and Pacific Tridacna lineages was already partly available. A distinctive T. maxima lineage and a newly rediscovered species (T. squamosina) had been reported from the Red Sea, but no phylogeographic information was then available from the western Indian Ocean (WIO). Hence our focus on Tridacna giant clams from that part of the tropical IWP.
During field work, several participants in this study – then working as separate teams – independently noticed giant clams initially identified as T. maxima but presenting somewhat distinctive features.
We noticed the sharply pointed triangular interstices between folds, and the remarkable emerald-green colour of the mantle edge.
Tridacna elongatissima from Etang Salé at Reunion Island. Photo credit: L. Bigot / Université de La Réunion.
Nucleotide sequences at the COI locus confirmed this giant clam was distinct from T. maxima, and from all other known Tridacna spp. then documented in public sequence databases. With an endemic species in the Red Sea (T. squamosina), two unverified rare species endemic to the Mascarene Basin (T. rosewateri and T. lorenzi), and now a new cryptic lineage in the WIO, we felt that we had an increasingly interesting subject to investigate. Our different teams merged efforts and datasets and pursued the phylogeographic work all together.
We managed to extract DNA from dried muscle tissue and ligament from the type material of T. rosewateri and from other specimens from the WIO region preserved in museum collections. Morphological and molecular analyses enabled us to identify the distinct Tridacna lineage present in the WIO as T. elongatissima, a long- forgotten species from Mozambique then synonymised with T. maxima, thereby adding a taxonomic hue to our primarily phylogeographic study. Meanwhile, T. lorenzi and T. rosewateri were found to be a single and same, distinct species.
This newly resurrected WIO-endemic Tridacna elongatissima turned out to be the sister species of T. squamosina! These two species had evolved independently in, respectively, the WIO and the Red Sea (or perhaps an adjacent northwestern Indian Ocean refuge), revealing a geographic barrier between the two regions. The T. elongatissima–T. squamosina pair was itself sister to T. rosewateri, highlighting this part of the world as an hotspot of endemism for giant clams. Lineage diversification patterns within the widespread T. maxima mirrored those of T. elongatissima, T. rosewateri and T. squamosina with two unrelated lineages in the WIO, one of which was sister to a third lineage endemic to the Red Sea. Thus, the same geographic barriers and speciation processes may have acted repeatedly at different periods in the Pleistocene.
We are aware, though, that no uniform explanation holds for the evolutionary history of species in the tropical IWP. At least we were able to refine our understanding of lineage diversification and endemism of Tridacna giant clams in the WIO and Red Sea region. Beyond the specific case of giant clams, our results emphasize the interest of sampling understudied regions of the tropical IWP, such as the WIO, to refine the evolutionary puzzle of this vast and complex geographic ensemble. Further investigations in the future may add to the story.
Written by: Philippe Borsa and Cécile Fauvelot – Researchers – Institut de recherche pour le développement (IRD), UMR ENTROPIE, France.
Antonia V. Mayr is a Postdoc at the University of Würzburg department of Animal Ecology and Tropical Biology. Her research is based in tropical mountain ecology and focuses on questions about how climate and land use change affect species, and how functional and phylogenetic diversity relate to changes in ecosystem services. Antonia provides background information on her recent work, which examines cavity-nesting pollinators and their natural antagonists on Mt. Kilimanjaro, Tanzania, where a large elevational gradient gives rise to different climates and ecosystems on a relatively small spatial scale.
(left) Antonia while doing her favourite work – fieldwork! She was also called mama nyuki (mother of the bees in Kiswahili) by her Tanzanian field assistants. (right) Antonia’s fieldwork was much nicer with often very interested friends. Here, they are checking occupied nests to see if they already hatched. If yes, Antonia took them to the field station, if no, she placed them back in the bucket.
Institution: University of Würzburg, Department of Animal Ecology and Tropical Biology
Current academic life stage: Postdoc
Research interests: My main research theme is tropical biology, especially tropical mountain ecology. Specifically, I am working on questions about how climate and land use changes affect species, but also functional and phylogenetic diversity and how this relates to changes in ecosystem services. I am fascinated by trophic interactions between plants, pollinators, predators and parasitoids and am very much interested in how to translate these findings into practical recommendations for conservation biology.
Current study system: Currently, I am studying cavity-nesting pollinators and their natural antagonists on Mt. Kilimanjaro, Tanzania. This is a very cool combination of study systems because the large elevational gradient of Mt. Kilimanjaro offers the possibility to study completely different climates and ecosystems on a relatively small spatial scale. Within few days, you can walk from the hot savanna through different mountain rainforests up to alpine ecosystems. Furthermore, the study of so-called trap nests enables us to investigate insects of different trophic levels, to collect data about functional and life-history traits and to study host-antagonist and food networks – which would be hardly possible to collect on this scale using only observational data.
A view of the Kilimanjaro summit from the field station in Nkweseko.
Recent paper in Journal of Biogeography: Mayr AV, Peters MK, Eardley CD, Renner ME, Röder J, Steffan-Dewenter I. 2020. Climate and food resources shape species richness and trophic interactions of cavity-nesting Hymenoptera. Journal of Biogeography 47: 854-865. https://doi.org/10.1111/jbi.13753
Motivation for the paper: Biologists have investigated the drivers of species richness for centuries. For insects, temperature, resource availability and top-down regulation as well as the impact of land use are considered to be important factors determining diversity. However, the relative importance of each of these factors is unknown. The steep climatic gradient and different land-use regimes at Mt. Kilimanjaro, together with the trap-nest system enabled us to simultaneously investigate the effects of temperature, resources, top-down control and land use on species richness of different trophic levels. With this paper, we hope to contribute to the discussion on drivers of insect species richness.
Key methodologies: We used trap nests, which are good bio-indicators for habitat quality and environmental changes, to collect and monitor cavity-nesting bees, wasps and their natural antagonists. To the best of our knowledge, this is the first trap-nest study in East Africa. We continuously monitored trap nests for 15 months, checking monthly for new nests and recently emerged insects. If you include the pre-experiment phase – in which we placed trap nests up to 4,240 m a.s.l and adapted the methodology – and the closing phase, our monitoring lasted 26 months in total. It is important to note that we hatched every nest on its respective study site. Normally, trap nests are collected, cut open and reared in the lab (in temperate regions during winter months) and/or brought back into nature after identification. In contrast, we did not take the nests to the research station before they hatched. The reason was that by working along an elevational gradient we would have changed the climatic conditions during the development of the inhabitants. This extra effort enabled us to collect year-round data about the occurrence of species, their development time and natural mortality, and link it to climate data.
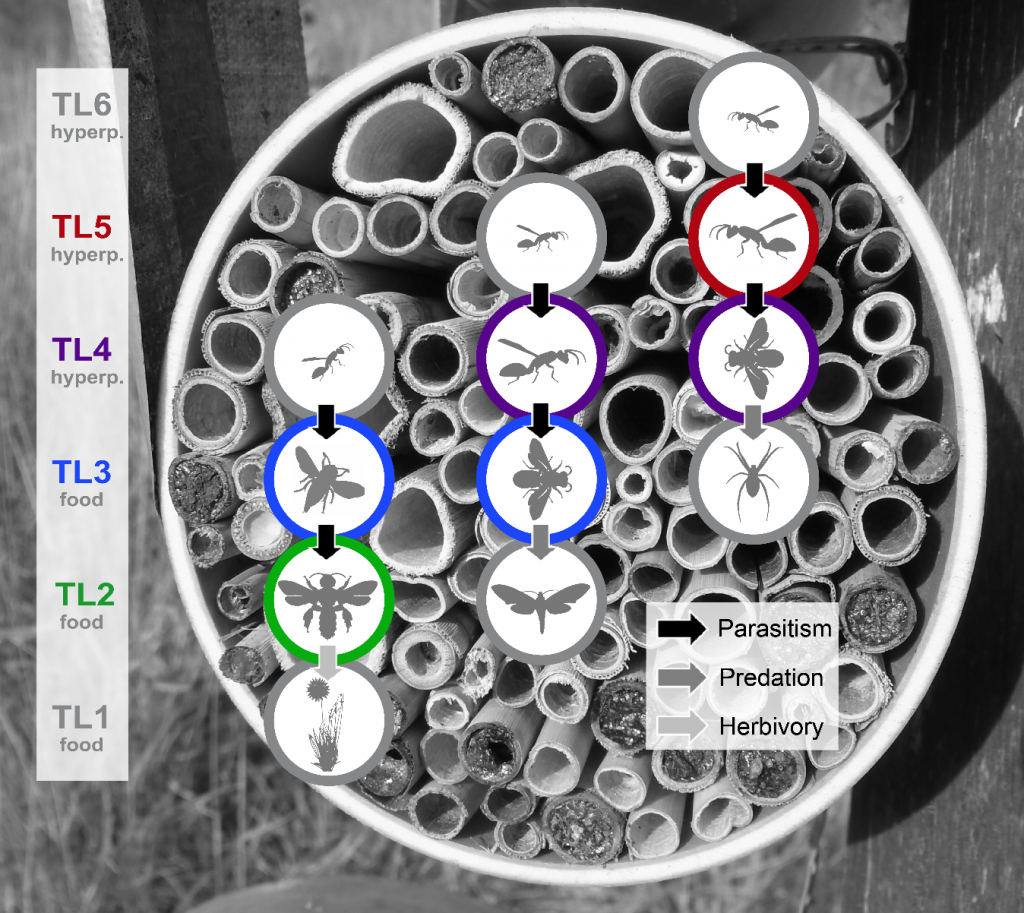
(top left) A pair of trap nests with the first version of the roofs, field assistant Jumanne, and Antonia. (top middle) An extra bucket attached to wooden poles with occupied nests waiting for hatching. Antonia closed the nests with metal nets instead of plastic nets because some bee species were able to cut their way out of it with their strong mandibles. (top right) An edited version of our roofs – the double-folded metal shields allowed the air to pass through and the nests underneath did not heat up anymore in a way that many larvae died. (bottom) Trap nests serve as a model system to investigate cavity-nesting communities and trophic interactions in different trophic levels. The picture shows the three different functional host groups which are subject to this study: bees (Apidae, Colletidae and Megachilidae), caterpillar-hunting wasps (Eumeninae) and spider-hunting wasps (Pompilidae, Sphecidae and Crabronidae), their respective natural antagonists, food resources and type of top-down effects. The trophic level affiliation is indicated by the colours and the colourful circles in green, blue, violet and red show the groups for which we analysed species richness patterns in this study.
Unexpected challenges: The biggest unexpected challenge was that a well-established method does not work well in super-hot and dry ecosystems, like the savannah during dry season. We used reed internodes (which are the sections of a reed stem) to artificially imitate natural cavities for cavity-nesting Hymenoptera. Bees in particular willingly nested inside the provided reed internodes. However, in our pre-experiment, many bee larvae died because they were either grilled by the heat and aridity or they drowned in their melted food made out of pollen and nectar. Our solution was to use double-folded metal sheets instead of plastic as roofs, allowing the air to pass through.
It was also surprising that there was a clear line at the border of the national park, above which no reed internodes inside the trap nests were occupied. Apparently, the relative humidity in the lower montane rainforest was too high and the temperatures too low to enable the occurrence of cavity-nesting Hymenoptera. Another lesson to be learned was that basically every material is valuable. It seems trivial, but it was very helpful to use torx screws to attach the roofs and trap nests to the wooden poles, as torx screwdrivers are still uncommon in Tanzania. This saved our materials from being taken away.
(left) A cut open brood cell. Here, the host, a megachilid bee did not emerge, but died for an unknown reason. (right) Pinning of bees and wasps for later identification. (bottom) Nest in which the megachilid bee mum used duct tape instead of leaf discs to construct the broodcells of her offspring.
Major result and contribution to the field: With 38 bee and 43 stem-nesting wasp morphospecies and 49 morphospecies of natural antagonists, we observed a very high diversity in roughly 4,050 nests. Our data suggest that temperature is a major driving factor for species richness patterns in bees, wasps and their natural antagonists. Furthermore, we found more trophic interactions in the warmer climates of the mountain, i.e. lowland ecosystems. By systematically analysing different trophic levels, we found that the importance of food resources increased for natural antagonists at higher trophic levels. Thereby, our study contributes to the discussion about the drivers of biodiversity along elevational gradients and provides novel insights into the relative importance of temperature, resources, trophic level and biotic interactions for bees, wasps and their natural antagonists.
What are the next steps? The next steps with this trap-nest dataset will be first, to understand seasonal changes in occurrence of species of different trophic levels. This is important because insect communities vary seasonally with related changes in climate and resource availability, strength of competition or pressure by natural antagonists, but are not well investigated in tropical mountain ecosystems. The second step is to investigate microclimatic effects on survival rates in savannah ecosystems, because upper thermal limits will first be reached in the lowland savannahs. Hence, it is useful to investigate how species cope with higher temperatures in partially warmer microhabitats to forecast effects of climate change. The third step is to investigate if traits are affected by land use, because traits may respond earlier to land use changes than species. Therefore, changes in traits might forecast changes in species compositions with increasing land use. The final step is to investigate if temperature and land use have an effect on host-parasitoid networks, which might be even more sensitive to environmental changes than species-richness patterns because they depend on very specialized species interactions.
If you could study any organism on Earth, what would it be and why? I was never totally restricted to one organism/group of organisms (before, I worked with ants). The better you get to know a group of organisms, the more fascinating they become. But I like diversity and enjoy not having only one model species. That is why I am very happy at the moment with bees and wasps, which show a high variety of forms and behaviour, e.g. in sociality and nesting. In addition, they inhabit very different ecosystems which not only makes fieldwork manifold, but offers the possibility to investigate adaptations to different environments, too.
Any other little gems you would like to share? Really impressive for me was the high diversity of nest-building behaviour. I was able to distinguish around 35 different types of nest closures – and this is only a study about cavity-nesting bees and wasps! What was outstanding for me, however, was that over time I discovered nests of leaf-cutter bees that used duct tape instead of leaves to build their broodcells and nest closures, with which I fixed broken trap nests. In the heat of the savannah, the duct tape stuck to my fingers so much that it was difficult for me to handle. How the bees cut the artificial pieces of leaves out of it without sticking to it is a complete mystery to me. Maybe bees are way more flexible and creative than we think they are…
(top) Diversity of nest closures, built out of soils, resins, plant leaves or fibres or secrets. (bottom) Antonia and her Tanzanian field assistants, who not only helped her to carry heavy luggage up the mountain, shoot trap nests high up the trees with a slingshot and check the trap nests, but they also contributed many creative and practical field solutions.
The above-ground diversity of plants in the Cape Floristic Region is legion. But what about arthropods? And what about below-ground?
Several years ago at the start of my postgraduate research training and career, I was part of a team of researchers from Stellenbosch University who were fascinated simply by how life works and how it is structured in the natural world. My interest in springtails was piqued on a research trip to sub-Antarctic Marion Island. Where my collaborators were absorbed by insects, I became fascinated with Collembola: how many species were there on the island? What were the differences between invasive and indigenous species, and what were these differences?
Image: Collembola such as the Seira species pictured above, show high turnover in the Cape Floristic Region. Photo by Amy Liu.
———————— FROM THE COVER: read the article on which this post is based … Janion-Scheepers C, Bengtsson J, Duffy, GA, Deharveng L, Leinaas HP, Chown SL (2020) High spatial turnover in springtails of the Cape Floristic Region. Journal of Biogeography, 47:1007–1018. https://doi.org/10.1111/jbi.13801 ————————
So I set about learning how to find out, with help from Steven Chown and Hans Petter Leinaas. Naturally, as soon as I had completed this work for my Masters thesis I asked another seemingly simple question: ‘How many species of springtails are in South Africa and what do they do?’ In the Cape Floristic Region, this question is almost inevitably also accompanied by speculation about whether any groups other than plants have also shown spectacular radiation. So my Ph.D. project and the rich collaboration with my co-authors were born, and the research that led to this paper.
The extraordinary diversity of plants across the Cape Floristic Region is characterized by considerable species turnover among sites. Studies so far on phytophagous insects show similarly high turnover, but their patterns are closely coupled to those of their hosts. Thus, if the mechanisms underlying high plant turnover are not unique to plants, similar patterns of turnover should also be seen in non-herbivorous arthropod groups. In this study, we tested this hypothesis for the Collembola fauna of the Fynbos biome. Thus, our research is testing major diversification ideas using an independent group to do so.
Diversity of environment and flora in the Cape Floristic Region
Most studies to date on alpha and beta diversity of the Cape Floristic Region have been on above-ground plants and invertebrates such as insects, showing there is extremely high turnover. This is the first study to investigate if this high turnover is also true for below-ground organisms. We foundthe alpha diversity of Fynbos Collembola assemblages is quite similar to that elsewhere, given energy availability and so on. By contrast, beta diversity is really high, and in particular considering the small distances among sites. That beta diversity is also characterized predominantly by species turnover. We were simply astounded by the high turnover we found for Collembola. Every site had new species, and some genera were extremely rich in diversity, with most species being undescribed. These patterns of unremarkable alpha diversity, but high turnover among sites are comparable to many Fynbos plant groups. The mechanisms giving rise to high beta diversity of the plants have probably led to high diversity in springtails independently of any host association. That’s an awesome finding!
This unexpected level of diversity in Collembola made clear one of the knottiest problems I had to overcome: Taxonomy. The world really needs more taxonomists. I have now had over a decade of training by arguably the world leader in Collembola taxonomy, Dr. Louis Deharveng from the Museum National d’Histoire Naturelle in Paris, France. This also led to more collaborations with other Collembola taxonomists specialising on certain families. So I am one more person that knows the systematics, at least partially, although we still need more. This problem is global and is a real barrier to finding out how our world works and what makes it work.
(This image, above, and below) Development and habitat change mean trouble for all of biodiversity in the Cape Floristic Region. The gems are both obvious and subtle, and all of them need conservation. Photos by Steven L. Chown.
Reflecting on the whole process, one of the major outcomes of the project has been an affirmation of my fascination in the drivers of turnover and the idea that some groups, other than plants and host-specific arthropods, have responded to the history and conditions of the Cape Floristic region to become highly diverse.
This project was also the start of several international collaborations with soil ecologists and taxonomists, and as mentioned above sparked my interest in taxonomy. This project also prompted our team to start using DNA barcoding to aid in species identification, and we now have a database of almost 2,500 sequences for Collembola in South Africa (www.boldsystems.org). These collaborations have led to several long-lasting partnerships and friendships.
Following this study, we now have several ongoing projects which have evolved from this one using Collembola as model organisms, sampling over a wider region, exploring their physiology, diet, taxonomic and molecular diversity, and their functional roles in the fynbos and other ecosystems in South Africa. Through this multidisciplinary approach, we hope to understand what the mechanistic drivers behind these shared above-below ground diversity patterns are. Our knowledge is certainly growing, although I still wonder: how many species of springtails are in South Africa and what do they do?
Written by: Charlene Janion-Scheepers – Lecturer, University of Cape Town, South Africa
Ida M. Mienna is a PhD student at Norwegian University of Science and Technology, having recently completed her master’s degree at Norwegian University of Science and Technology. Her current work focuses on understanding the spatial ecology of the forest-tundra ecotone in Fennoscandia, especially through investigating drivers of change and establishing a high resolution monitoring system using drone imagery and airborne laser scanning. Ida discusses her recent work on the flora of Norway, which explores evolutionary signals in plant distributions using DNA from herbarium specimens.
Although Ida’s study did not require any data collection in the field, she went on many trips to understand the ecology and spatial distribution of vascular plants in Norway. Here Ida is posing with the orchid Nigritella nigra, a priority species in Norway, at its northernmost known locality in the world.
Institution: Faculty of Environmental Sciences and Natural Resource Management, Norwegian University of Life Sciences (NMBU); Department of Natural History, NTNU University museum, Norwegian University of Science and Technology (NTNU)
Current academic life stage: PhD
Research interests: In general, I am really interested in anything that has to do with spatial ecology. I guess that is why I find research within community ecology and plant biodiversity distributions exciting, as these have clear spatial patterns in them. For my master’s in biology I was introduced to the spatial phylogenetics approach that I used in this study, which combines spatial ecology and evolutionary history. For my PhD, I am currently going into depth on how one can use remote sensing techniques to study changes in spatial distributions of communities. So, to summarise, I am interested in research that involves field, lab or computer work – it does not matter, as long as it has a spatial component to it!
Current study system: I am working with the forest-tundra ecotone, also known as the treeline. In general, the treeline is expected to increase in elevation with rising temperatures, but the extent to which this happens and what other factors may affect its dynamics are still not well understood. Thus, I am looking into drivers like herbivory density in addition to temperature. Establishing a high-resolution monitoring system also is important as the treeline is not changing rapidly; for this I am exploring the use of remote sensing, i.e. drone imagery and airborne laser scanning.
Recent paper in Journal of Biogeography: Mienna, I.M., Speed, J.D.M., Bendiksby, M., Thornhill, A.H., Mishler, B.D., Martin, M.D. 2020. Differential patterns of floristic phylogenetic diversity across a postglacial landscape. Journal of Biogeography 47: 915-926. https://doi.org/10.1111/jbi.13789
Motivation for the paper: Exploring spatial distributions of biodiversity is important so that we know where we should focus our conservation efforts, but often we focus on areas where we have the most species. Research also often focuses on areas already known to be species rich and that have long evolutionary histories. In this study we wanted to explore the flora of Norway, which consists of plants that had to immigrate to the area after the Fennoscandian Ice sheet retreated 22-9.7 thousand years ago. We wanted to discover if there were any evolutionary signals in the plants’ distributions and if we found patterns that you normally would not find by looking at species diversity alone (i.e. potential places with concentrations of relatively young or old species).
Key methodologies: To study evolutionary patterns, we needed to make a phylogeny of the native Norwegian flora, which is—as far as we know—the first time this has been done. Most of the DNA sequences were downloaded from various sequence databases, but not all species had available data. Instead of sampling our own data from the field, we collected DNA samples from herbarium specimens, which was really nice as I worked at a museum with all the resources needed around me. Information about plant distributions were downloaded from GBIF, so for these data we did not need to go into the field either. The phylogeny and the distribution data were then combined to study the evolutionary patterns. We also modelled different diversity patterns to see if predictors like temperature, precipitation and time since the area was covered by ice affected the spatial patterns.
(left) The vascular plant Pedicularis hirsuta, which in Norway can only be found in the northern mountains. This species is regarded as near-threatened and is expected to decline in numbers with increasing temperatures. (right) To estimate the phylogenetic diversity of vascular plants across Norway, a phylogeny based on DNA sequences was made. Many of the sequences came from DNA samples sampled from herbarium specimens, like this one.
Unexpected challenges: Working with DNA from herbarium specimens is not as easy as working with fresh samples, and there were some struggles in the lab getting the PCR to work. However, having helpful and experienced supervisors is always good and we managed to get (almost) all three gene sequences for all species. Working with big data is also not that straightforward as many of the data have errors (e.g. a downloaded DNA sequence was actually from another species or the species occurrence was inaccurate) and finding ways to remove as many of the errors as possible while not removing valid data can be difficult. However, we believe that the methods we used to avoid these errors worked quite well in the end.
Major result and contribution to the field: We found spatial patterns that had not been seen in Norway, but also patterns that were concordant with previous studies. The cold and dry mountainous areas had significantly lower phylogenetic diversity than the rest of the country, suggesting that the species there were relatively closely related to each other. The warmer and wetter coastal areas had significantly higher phylogenetic diversity, suggesting phylogenetic overdispersion possibly due to competitive exclusion. We also found that in the northern parts, many areas have quite young plants (i.e. recently evolved). As this pattern also was mainly found in dry and cold climates, we suggest that recent speciation due to stressful environments may be the factor behind these patterns. All these results show an evolutionary signal in the distribution of the vascular flora of Norway.
What are the next steps? Currently, we are expanding the study area to include Sweden and Finland in addition to Norway, representing the native flora of the Fennoscandian Peninsula. This area, like Norway, was covered by the Fennoscandian Ice Sheet, meaning that all plants in the area came after the ice melted. We will do the same analyses as in our previous paper, but also see if the distributions have changed in the last 100 years and if their future distributions will be affected by climate change. In addition, we are interested in seeing if the protected areas actually contain most of the phylogenetic and species diversity.
If you could study any organism on Earth, what would it be and why? I have never really studied one organism alone, but rather communities. Funnily enough, considering that I chose to study terrestrial plants, I have always found colourful marine organisms like nudibranchs and polychaetes fascinating. Unfortunately, I have a little bit of thalassophobia, so I will stay on land studying colourful plants for now.
Any other little gems you would like to share? During my master’s in biology when I started working on this paper, I did not, as mentioned above, do any field work related to my research. However, I had a motivation to learn more about my study organisms, which led me to go hiking in the forests and mountains to learn the names of the plants and their ecology. I think this is when I realised that I needed to do a PhD, as just doing one simple task was not enough and I needed to explore further to understand what I did not understand.
Norway only has about 1250 native vascular plant species, but the country stretches widely both in latitude (50-71° N) and longitude (4-32° E) and has a large topographic variation (0-2469 m a.s.l.), making it a very interesting study area. This picture shows a south-facing slope with high, relatively vascular plant diversity.
How to tease apart the mechanisms behind patterns of diversity in the wild.
A major challenge in ecology and evolution is to unravel the processes that generate and maintain the uneven distribution of life on Earth. A common approach is to regress species richness, the number of species counted at a place, on variables representing contemporary and historical environmental conditions. However, numerous hypotheses for richness gradients implicitly or explicitly make predictions about mechanisms acting on how species occupy niche space, the degree of ecological differentiation among species, and therefore to trait dispersion within assemblages, which cannot be addressed using data on species richness alone. For instance, contemporary hypotheses predict that areas of higher primary productivity have a larger available niche space that might be filled with more species. Moreover, historical hypotheses predict that areas that remained stable through geological time (i.e., over millennia) suffered less from extinctions and allowed the accumulation of more species, lineages and trait diversity over longer periods.
Image: A view of the Australian Wet Tropics landscape taken from the Thornton Peak. Photo by: Stephen Zozaya..
———————— FROM THE COVER: read the article on which this post is based … Oliveira, BF, Flenniken, JM, Guralnick, RP, Williams, SE, Scheffers, BR. (2020) Historical environmental stability drives discordant niche filling dynamics across phylogenetic scales. J Biogeogr. 47: 807–816. https://doi.org/10.1111/jbi.13798 ————————
To test these predictions, we focused our study in the Australian Wet Tropics (AWT), a region that has undergone significant historical habitat shifts, with rain forests expanding and contracting over the Quaternary period. Here, we investigated how spatial patterns of niche space occupancy conform to current environmental conditions (Net Primary Productivity, NPP) and historical stability of environmental conditions since the Last Glacial Maximum (LGM, ~21 kyr BP). Following Robert Helmer MacArthur in his seminal paper (MacArthur, 1965), we estimated two contrasting ways for species to occupy niche space: packing and expansion. Niche packing occurs when increasing richness leads to a denser packing of species into a niche space, whereas niche expansion occurs when species diversify into novel regions of niche space resulting in increased richness. We used datasets of unusually high phylogenetic scope, including all known vertebrates (mammals, birds, amphibians, and reptiles) occurring in the AWT, species distribution data derived from 20 years of standardized surveys, and trait data of measurements taken preferentially from specimens occurring at the region.
A series of previous studies conducted at the AWT showed that historical environmental fluctuations played an important role in shaping current biodiversity patterns in the region, including species distribution ranges, richness patterns and genetic structure of populations. Our study indicates that historical processes also determined dynamics of niche space filling and occupancy. Across multiple taxonomic groups, stable areas tend to be “expansive”, with niche dynamics driving assemblages of higher functional diversity than expected by chance. Although current environmental productivity and historical stability are linked, productive regions cannot support high biological diversity if they are located in unstable areas. As a result, the net effect of NPP on richness and niche occupancy patterns was offset by a stronger effect of historical environmental instability, reiterating the importance of historical processes for the generation of current diversity gradients.
Our broad phylogenetic sampling is of particular utility for examining inter-clade interactions in the filling of niche space. We identified discordant niche occupancy patterns across analyses of differing phylogenetic scale (e.g., mammals versus all vertebrates). At lower phylogenetic scales (i.e., within a given constituent vertebrate group), increases in species richness lead to niche differentiation, a characteristic of niche expansion. In contrast, at higher phylogenetic scales (i.e., the scale of vertebrates), increases in richness increases the sharing of trait space by species from different constituent classes, a characteristic of niche packing. These seemingly contradictory results suggest that dynamics of niche space occupancy depend on phylogenetic scale. Further cross-phylogenetic scale analyses may allow researchers to have a more complete picture of the processes that generate and maintain the diversity of life.
Written by: Brunno F. Oliveira – Postdoctoral Researcher, Department of Wildlife Ecology & Conservation, University of Florida/IFAS, Gainesville, Florida, USA Robert P. Guralnick – Curator of Biodiversity Informatics, Florida Museum of Natural History, University of Florida, Gainesville, Florida, USA Stephen E. Williams – Professor, Centre for Tropical Environmental & Sustainability Science, James Cook University, Townsville, Australia Brett R. Scheffers – Assistant Professor, Department of Wildlife Ecology & Conservation, University of Florida/IFAS, Gainesville, Florida, USA
Zach Carter is a PhD student at the University of Auckland School of Biological Sciences, focusing on invasion biology, island ecosystems, and physical geography. He is currently studying landscape-scale eradication of invasive mammals of the New Zealand archipelago, including rats, mustelids, and the common brushtail possum. Zach shares the motivations and challenges behind his recent publication in the Journal of Biogeography, which aims to improve the understanding of reinvasion potential and to identify the greatest threats to the long-term eradication of invasive species.
(left) Zach sexing a kiore (Rattus exulans) on Slipper Island off of New Zealand’s Coromandel Peninsula. (right) Zach reluctantly holding a deceased eel (Anguilla sp.) on Motukawanui, an island off of New Zealand’s Northland coast that was featured in his recent article.
Institution: University of Auckland, School of Biological Sciences
Current academic life stage: PhD
Research interests: I am interested in prioritising landscape-scale eradication projects of invasive species to achieve desirable conservation outcomes. My work has a strong statistical modelling and analytical component, focusing primarily on invasion biology, island ecosystems, and physical geography.
Current study system: I am currently studying invasive mammals of the New Zealand archipelago, including rats (Rattus rattus, R. norvegicus, R. exulans), mustelids (Mustela erminea, M. furo, M. nivalis) and common brushtail possum (Trichosurus vulpecula). New Zealand’s endemic biota evolved in the absence of terrestrial mammals, making them highly susceptible to mammalian predation. As a result, the species I study constitute a grave biological threat even though they are relatively unassuming (and even cute). Understanding how rats, mustelids and possum affect their surroundings, and how their negative impacts can be reversed, is particularly interesting.
Recent paper in Journal of Biogeography: Carter ZT, Perry GLW, Russell JC. (2020) Determining the underlying structure of insular isolation measures. Journal of Biogeography 47:955–967. DOI: 10.1111/jbi.13778
Motivation for the paper: Our motivation for this study was to better understand reinvasion potential; reinvasion represents the greatest threat to long-term eradication success. Rodents and stoats readily hitchhike to islands as seafaring stowaways or disperse naturally by swimming between islands. As a result, an island’s reinvasion risk is directly correlated with its level of geographical isolation. However, quantifying insular isolation is not necessarily a straightforward process: there are many different metrics describing isolation and it can be difficult to ascertain which metrics are necessary for quantification. We thought resolving this issue would make for a terrific study. Because isolation is such an important aspect of biology, we made a concerted effort to make our findings applicable and accessible across disciplines.
(left) Common brushtail possum (Trichosurus vulpecula) and (right) the Polynesian rat (kiore, Rattus exulans). Both species have had a significant impact on New Zealand’s biota through predation and resource competition. New Zealand’s endemic species evolved in the absence of terrestrial mammals and, as such, are highly susceptible to their threats.
Key methodologies: We conducted a principal components analysis (PCA) on 16 different measures frequently used to describe insular isolation, including Euclidean-based distance metrics, landscape connectivity measures derived from least-cost and circuit theory modelling, stepping stones, and buffer metrics, among others. PCA simply accounts for redundancy within a set of input variables; variables that describe a similar thing will cluster (or ‘load’) together. We used these clustered components (the ‘principal components’ or PCs) to understand what variables may be necessary for measuring insular isolation.
Unexpected challenges: Interestingly, we found that a simple Euclidean distance measure describes insular isolation comparably to graph-theoretic and least-cost connectivity metrics. Connectivity metrics are incredibly useful for understanding aspects of a heterogeneous environment, but are apparently limited in areas that are homogenous and inhospitable (such as an ocean body to a terrestrial mammal). Moreover, calculating a Euclidean distance is straightforward because it is simply the straight-line distance between two locations. Connectivity metrics, on the other hand, are often computationally intensive. We were required to calculate our least-cost paths, resistance distances, and their derived metrics using a high performance cluster computer. These calculations took months of continuous computing due to the scale of our analysis. In the end, we found that a simple 5-minute calculation was comparable to one taking hundreds of hours to complete. I had to take a small vacation after I completed this paper in order to keep my sanity in check.
Major result and contribution to the field: The basis of a robust description of insular isolation comes from three independent PCA components, including distance from the mainland source to the focal island (PC1), stepping stones available along the dispersal pathway (PC2), and the focal island’s position within the landscape (PC3). Importantly, these PCs are relatively easy to calculate. PC1 can be measured using a simple Euclidean distance measurement, PC2 by counting the number of stepping stone islands available along a dispersal pathway, and PC3 by measuring the focal island’s size. The parsimony and simplicity afforded by these measures should facilitate application across disciplines.
What are the next steps? We make no claims regarding the ecological importance of the identified principal components. The PCA cannot tell us which measures are most important, only what measures are necessary for describing isolation. As such, we place all three identified PCs on equal footing. A significant next step in this research would be to identify the relative importance of each component. Doing so would allow for creation of an equation describing insular isolation with a single metric. Our paper only focused on small mammalian dispersers, though. It is very likely the components describing isolation would look entirely different for other dispersers (particularly those that are well-equipped for long-distance dispersal).
If you could study any organism on Earth, what would it be and why? I feel incredibly lucky to be studying New Zealand’s invasive species. In 2012, the New Zealand government championed the Predator Free 2050 programme, an initiative to extirpate rats, stoats, and possum from the entirety of the country by 2050. This programme is a first of its kind and I couldn’t be happier to be studying the organisms and systems that I am. I hope to continue my work on these invasive species even after completion of my PhD.
Any other little gems you would like to share? I have taken quite a meandering path to get to where I am today: my BSc and MSc are both in engineering. Studying complex biological systems was initially difficult to grapple with at the beginning of my PhD but I am finding my engineering background has been exceedingly helpful. The time I’ve spent working as an engineer has given me a fresh perspective to problem solving that I wouldn’t have had otherwise. I am proof that having an initial lack of direction can turn out to be a beneficial career move!
Islands at the northern end of the Cavalli Islands. Panaki Island (identified under the arrow) was used as the example focal island in our recent paper. The Cavalli’s are a great example of how stepping stone islands facilitate dispersal of terrestrial biota.
How niche conservatism in colonizing and sedentary species shape a latitudinal gradient.
Niche conservatism has often been used as an elegant explanation for why there are more species in the tropics—i.e. most taxa originated in the tropics, had more time to diversify therein, and tended to retain ancestral climatic affinities making range shifts out of the tropics less likely. However, researchers have rarely referred to niche conservatism to explain why some taxa exhibit inverse patterns of increasing diversity towards the poles. We wondered whether niche conservatism would also make a valid explanation for taxa exhibiting inverse richness gradients. In other words, would clades originated under extra-tropical climates still be more diverse outside the tropics?
Image: Illustration of examples of clades that participated in the Biotic Exchanges through Bering in either direction, from the Palaearctic to the Nearctic and from the Nearctic to the Palaearctic. The species belonging to these clades are classified as ‘colonizers’ and assumed to be adapted to cooler temperatures to some extent.
———————— Read the Full (open) Access EDITORS’ CHOICE article on which this post is based: Morales‐Castilla, I, Davies, TJ, Rodríguez, MÁ. (2019) Historical contingency, niche conservatism and the tendency for some taxa to be more diverse towards the poles. Journal of Biogeography 47:783– 794. https://doi.org/10.1111/jbi.13725 ————————
While it is hard, if at all possible, to pinpoint the geographic origin of current clades, palaeontological records provide highly valuable information. For example, the fossil record contains information about past distributions and historical dispersal events. We thought of the mammalian biotic exchanges through the Bering Strait as a natural experiment allowing classification of species into colonizers—i.e. current species descending from ancestors that crossed Bering—and sedentaries—i.e. species whose ancestors did not cross Bering. Assuming that colonizers would have been adapted to cooler climates, we tested if, despite the long timespan since these dispersal events took place, colonizers would still distribute preferably over the extra-tropics.
The answer is yes, while sedentaries follow the classical Latitudinal Diversity Gradient (LDG) –i.e. they are more diverse toward lower latitudes– and show negative temperature-richness relationships, colonizers exhibit the reverse latitudinal trend increasing in diversity with cold. What was surprising to us was that, being classified as a colonizer according to the (admittedly sparse) fossil record would better explain climate-richness relationships than usual suspects in macroecology such as body size or range size. Altogether, our results step towards reconciling views explaining the LDG based alone in climatic determinism, evolutionary rates, or tropical conservatism. Cold adapted ancestors were able to cross Bering, but their descendants had a harder time moving towards tropical latitudes because extra-tropical niches are conserved too.
Our paper confirms that historical contingency, dispersal and niche conservatism, matter when it comes to understanding contemporary diversity gradients. This would have implications for species currently adapted to cold climates, in a context where regions with cooler climates are shrinking. More importantly, our findings lead us to ask new questions such as, are historical dispersal events as important for plant and animal taxa other than mammals? Are there physiological traits able to explain why some clades and not others participated in biotic exchanges? How much further can we incorporate palaeontological findings into the biogeographic agenda? How long back in time, can we track the imprints of history and evolution into the contemporary distribution of biodiversity? Would any of this information be useful to hindcasts or forecasts of species distributions? We are excited to tackle some of these questions and hope that fellow biogeographers will join us in this fascinating research venue.
Written by: Ignacio Morales-Castilla Postdoctoral Fellow, Global Change Ecology and Evolution (GloCEE), Department of Life Sciences, University of Alcalá, Spain
x Differing diversity-climate relationships—i.e. species richness vs. temperature—between sedentary species, which conform to the LDG and show positive slopes and higher species richness towards the tropics (yellow boxplot and ribbon); and colonizers, which show a higher proportion of negative slopes and higher species richness across temperate and cooler climates.
Letícia Soares is a postdoc at the Advanced Facility for Avian Research, University of Western Ontario, and is interested in disease ecology and evolution. In her recently published paper in Journal of Biogeography, Letícia studied the malarial parasites in North American migratory bird species in their Hispaniola over-wintering range. She shares her findings from her temporally stratified collections and the differences between migrant and co-occurring resident bird species.
Letícia during field work in Bruce Peninsula (Canada), working on yellow-rumped warblers during their fall migration.
Institute. Advanced Facility for Avian Research, University of Western Ontario
Academic life stage. Postdoc
Major research themes. Disease Ecology & Evolution, Biogeography
Current study system. Most of my current research focuses on determining the drivers of geographical and temporal variation in the occurrence of malarial parasites in birds (order Haemosporida). In birds, malarial parasites have a relevant regulatory role in populations, as infections can reduce the reproductive success, life span, and survival of hosts. Moreover, bird malaria is biologically similar and evolutionary related to human malaria, making this the perfect study system to understand the ecology and evolution of malaria parasites in natural populations.
Recent paper in Journal of Biogeography. Soares L., Latta S. and Ricklefs R. (2020) Neotropical migratory and resident birds occurring in sympatry during winter have distinct haemosporidian parasite assemblages. Journal of Biogeography, 47(3):748–759. https://doi.org/10.1111/jbi.13760
Motivation behind this research. Neotropical migratory birds have a life history strategy that most would envy: they spend their summers breeding in North America, and when days become shorter and food supplies start to diminish, they make their way southwards to their wintering grounds, where migrants will spend up to five months co-occurring with year-round resident bird species. Most migratory birds are infected with malarial parasites prior to travelling to wintering areas, but we do not know the effects of these parasites on host survival during North Hemisphere winters. We additionally have very little understanding of the fate of parasitic infections during wintering. For migratory birds, infections likely confer negative effects that can carry over to future breeding and migratory seasons. Hence, we wanted to quantify how often birds that reside year-round in the Tropics exchange parasite strains with migratory birds. We also wanted to assess the potential of migratory birds as vehicles for parasite dispersal across large geographical scales. For example, picture a Cape May warbler during the breeding season in Maine, USA: this bird gets bit by a mosquito that transmits a malarial strain to it. When this bird travels to Hispaniola for the winter, it will co-occur with birds that reside there year-round. If this particular Cape May warbler brings this North American malarial strain to Hispaniola, it has the potential of transmitting that parasite to local resident species via mosquito vectors.
Two common migratory species caught while mistnetting in the Caribbean. (left) Cape May Warbler. (right) Black and White Warbler.
Methodology. Consistent sampling was important in collecting high quality data. This was only possible thanks to Dr. Steven Latta and collaborators in the Dominican Republic. Capturing, banding, and taking blood samples from birds in remote areas is no easy task and always deserves to be acknowledged and appreciated. It was a game changer to be able to demonstrate that migratory birds are rarely detected with infections across multiple years, even on years of high prevalence among resident species. Before this study, we worked with resident bird species in more than 20 islands of the West Indies, demonstrating that infection prevalence exhibits dynamic fluctuations on short time frames (2–3 years). Thus, our sampling allowed us to demonstrate that even though migratory and resident birds might face similar ecological conditions in Hispaniola, migrant populations seem refractory to what happens to resident populations, at least when it comes to malarial parasites.
Unexpected outcomes. Our data show a very strong and multi-year pattern of low prevalence of malarial infections among migratory birds, even in years when co-occurring populations of resident birds have infection prevalence as high as 78%. My a priori expectation was that migratory birds would be found infected with parasite lineages that commonly occur in the wintering grounds, which was not the case. Elaborating biologically meaningful hypotheses to explain these findings was quite challenging, given that very little is known about the winter ecology of malarial parasites in migratory and resident bird populations, and the information on vector biology for this system is also scarce.
Major results. We have demonstrated that during midwinter on Hispaniola, populations of 14 over-wintering migratory bird species are rarely infected with malarial parasites. In all five sampled years (2001, 2002, 2014, 2015, and 2017), we found that, on average, 27% of 1,780 resident birds were infected with malarial parasites, but infections were detected in only 2.8% of 901 over-wintering migratory individuals. While previous studies provided the valuable insight that parasite dispersal by migratory birds is limited, they were focused on determining the prevalence of infections at breeding, migration stop-over and wintering areas of single species of migratory birds. Hence, prior studies lacked multi-year sampling and data on other resident bird species that co-occur with these migrants during different phases of their life-cycles. What makes our study stand out is that we expanded sampling from populations to communities to focused on assemblages of migratory and resident birds occurring in sympatry during North Hemisphere winters, with considerable temporal replication. We inferred that malarial parasite seldom use migratory birds as vehicles for dispersal, and that migratory birds rarely exchange parasite strains with resident birds of Hispaniola.
(left) Broad billed tody, a Hispaniola endemic species. (right) Haemosporidian parasites have complex life cycles. Within the vertebrate hosts, these parasites replicate clonally and produce transmission stages (highlighted by the arrows) that infect red blood cells.
Next steps. Like human malarial parasites, bird malarial parasites are transmitted by blood-feeding insects. To fully understand what determines the ability of a vector-transmitted parasite to disperse and establish transmission cycles in a novel geographic region, we need to describe the communities of insects that prefer to feed on bird blood, and determine whether these blood feeding insects are suitable vectors for the parasites. Diminished vector populations during North Hemisphere winters is among the hypotheses we proposed to explain the low frequency of parasite exchange between resident and migratory birds. We also tend to intuitively think of blood-feeding insects as being opportunistic feeders, when in reality many do present feeding preferences. Thus, we cannot rule out the possibility that insect vectors in Hispaniola are not suitable vectors for North American parasite strains, or that these insects are just not fond of drinking blood from migratory birds.
If you study any organism on Earth, what would it be and why? I would love to study bird species that seem refractory to commonly occurring parasites. For example, malaria parasites have been found infecting most bird species that have been sampled, with the exception of shorebirds. Why shorebirds are rarely detected with malaria infections is not understood, but it is possible that shorebirds are incompatible hosts for these parasites. Most shorebirds that breed at high latitudes in the Northern Hemisphere undertake long-distance migration to wintering areas in South America. Infections likely increase the costs of migration, and, over time, natural selection would have favored individuals with effective immune response against infections that increase the costs and affect the energetic budget of migration. Understanding the mechanisms regulating disease susceptibility and resistance in birds can inform research into human malaria, as many strategies of host exploitation by the parasite and immune response by the host are similar across the avian and mammalian malaria parasite systems.
Anything else you would like to share? In my capacity as a postdoc at the University of Western Ontario, I am experimentally determining the effects of malaria parasites on the migratory performance of yellow-rumped warblers. I am using experimental infection and simulated migration in a wind tunnel to determine whether malaria parasites affect migratory flight performance by integrating metrics of flight behavior, infection intensity, and costs of infection with a direct association to host longevity and physiology. The results from this experiment could help us explain the low prevalence of malaria infections in migratory birds during wintering. Migration is physiologically strenuous for the birds, and we hypothesize that oxidative stress resulting from endurance exercise during migration could have a negative effect on within-host parasite survival.
Grasses are extremely cosmopolitan in distribution and comprise one of the largest biomes on Earth. They are important carbon stores, account for large amounts of terrestrial primary productivity, are home to much of the worlds mammalian diversity and contribute to the livelihoods of an estimated one fifth of the world population. Grasslands are culturally, economically and environmentally important and there is an increasing need to predict how these ecosystems are likely to respond to global anthropogenic change. In order to do so we must first understand how they are assembled, their structure and how they are maintained.
Read the EDITORS’ CHOICE article on which this post is based: Jardine, EC, Thomas, GH, Forrestel, EJ, Lehmann, CER, Osborne, CP. (2020) The global distribution of grass functional traits within grassy biomes. Journal of Biogeography 47:553– 565. https://doi.org/10.1111/jbi.13764
Image (top): Sudanian tropical savanna with clumps of Andropogon gayannus courtesy of Marco Schmidt (Wikicommons)
(Left) Spinifex savanna in MacDonnell Ranges, Central Australia courtesy of Thomas Schoch. (Right) An example of short Steppe grassland between Morin and Erdenet, Mongolia, image courtesy of Jamie Ingram.
Trait based approaches to ecology make generalizations regardless of species identity, to predict individual plant responses to environmental variation which can scale up to explain the processes that are responsible for community assembly, ecosystem function and global vegetation dynamics. In this piece of work, we wanted to test whether predictions made by community ecological theory are more broadly applicable at larger scales, asking whether traits that explain performance at community scale have the same relationships to environment over global scales.
Studying the global relationships of traits with environment brings with it some challenges. Firstly, identifying which of the more than 11,000 species of grass we would sample and also finding data that document the distributions of these species across the globe. The precursor to this paper was a global map of the worlds grassy biomes which had been produced by a NESCENT working group. Using the descriptions and metadata from vegetation maps, colleagues had been able to identify different grassy vegetation types and the dominant grass species which characterise these for global grassy biomes. This map became the basis of the species sampling for this work.
We considered grasslands to include vegetation with an open canopy and continuous ground layer as shown in this photo taken near Grahamstown, South Africa.
The second problem was accessing plants from which to take trait measurements. Functional traits are usually measured from fresh material, however it was simply not within the scope of this project to collect fresh, wild material from all across the world. To overcome this problem, we developed methods for measuring functional traits from herbarium specimens. We were fortunate enough to be given access to the collection at the Royal Botanical Gardens, Kew. Here we found grasses that had been collected from all continents, spanning a global soil and climatic gradient and from all of the major and most minor grass phylogenetic clades. Herbaria document a staggering amount of the worlds flora and have long been important repositories for the study of taxonomy and geographic distributions. This work really exemplifies the importance of herbarium collections as a source of ecological and evolutionary information. Within the collection at Kew, we were able to sample around one quarter of the worlds globally dominant grass species and this was mostly constrained by time and not the collection.
Through this project we discovered weak relationships between commonly measured economic traits (specific leaf area, leaf tensile strength) and also size related traits with contemporary environment at global scales. We showed that evolutionary history explains more trait variation than contemporary climate and that large amounts of trait variation occurs at small scales (i.e. within rather than between vegetation types). This information is important for modellers interested in how ecosystems will respond to climate change.
Much of trait based ecology has to date focussed upon leaf economics, probably in part because these traits are quick and simple to measure. Our work indicates that some of the most commonly measured functional traits may not be able to improve our predictions of how species and ecosystems respond to global change. When relationships between economic traits and environment are not observed, this does not however mean that trait relationships do not exist; another axis of trait variation may be important in explaining contrasting species distributions. The findings of this study have prompted us to investigate other axes of trait variation beyond the leaf economic spectrum, including physiological and also underground traits and how these relate to environmental variation.
Ryan Burner is currently a postdoc at the Norwegian University of Life Sciences. He is a community ecologist with an interest in the birds and insects harboured in tropical ecosystems. His recent work in the Journal of Biogeography has investigated the range limits of birds inhabiting Bornean mountains. Ryan shares his findings on temperature gradients and the “fine-tuning” effects of competition in structuring these montane bird communities from South East Asian.
(left) Ryan Burner with a Banded Broadbill (Eurylaimus harterti) mist-netted in Danum Valley, Sabah, Malaysian Borneo. (right) Oriental Paradise-flycatcher (Terpsiphone affinis) mist-netted on Mt. Mulu, Sarawak.
Institute. Norwegian University of Life Sciences (NMBU)
Academic life stage. Postdoc
Research interests. Community ecology, Tropical ecology, Ornithology, Entomology, Bayesian modeling.
Current study system. Much of my work takes place in Borneo in Southeast Asia – one of the largest islands in the world – with a focus on birds. Borneo has more endemic bird species than any of the surrounding islands and also has some of the most intact remaining tropical forest. It is also a place with a lot of conservation challenges, however, and more forest is being cut down every day to make forest-derived products and clear space for plantations. The biodiversity in the tropics is incredible, and the basic life history of so many species unknown. For example, my co-authors and I have been able to describe the nests of several bird species for the first time, and many remain to be discovered. Louisiana State University (where I completed my PhD) and University of Montana and The Evergreen State College (homes of several collaborators) are leading the way in understanding the birds of this fascinating region!
Recent paper in Journal of Biogeography. Burner, R., A. Boyce, D. Bernasconi, A. Styring, S. Shakya, C. Boer, M. Rahman, T. Martin, F. Sheldon. 2020. Biotic interactions and temperature gradients contribute to differences in bird range limits on Bornean mountains. Journal of Biogeography 47: 760-771. https://doi.org/10.1111/jbi.13784
Research motivation. As ecologists, we are interested in why species occur where they do. This is particularly interesting on tropical mountains, where you can walk uphill for a few hours and pass into almost entirely new bird communities. What stops all these species from living farther down (or up) than they do? Is it climate? Habitat? Competition with other species? This is particularly important as we think about climate change. Will some of these species change their ranges, or lose their homes entirely as they are forced onto tiny mountain tops?
One way to answer this is to compare the current distributions of a group of species to their distributions in the past and see whether any differences are correlated to changes in the habitat or climate. The problem is that there are few historic data sources of sufficient quality to do this. What we did instead was compare the ranges and relative abundance of a group of over 100 bird species on three different mountains in Borneo. These mountains differed in their habitat, climate, and the abundances of potentially competing bird species, letting us test which of these factors is most important.
Key methodologies. When looking for the effects of competition, ecologists often test whether a given species is less common when another similar (but potentially competing species) is more abundant. If so, this is taken as evidence that the two species may compete. Theory predicts, though, that in many cases competition won’t just occur between pairs of species but rather will be among larger groups of species. But it is harder to test for this ‘diffuse’ competition. What we did was add together the abundances of entire groups of potentially competing species – basically species that ate similar foods in similar ways – and test whether these total abundances could be used to predict the abundances of other species.
Unexpected challenges. I love working in remote areas and it is a priority in my research, but it does have its challenges. There were times during the fieldwork for this project, hiking steep slopes in the rain before sunrise, that we questioned the sanity of what we were doing. But in exchange for a bit of discomfort there are incredible rewards – the huge trees, beautiful streams, strange sights and sounds.
Another challenge was a bit more mundane – listening to and transcribing many hours of recordings after returning home to identify difficult bird species. But even this can take you back to the forest. Listen to this Crimson-headed Partridge (Haematortyx sanguiniceps).
(left) A typical small stream from the steep montane forest of Borneo. Much of Borneo receives more than 3 m of rainfall per year. (right) Poison-tipped blowgun darts carried by a local hunter near Mt. Mulu. Several Penan villages are located near the park, and they are expert at living in the forest.
Major results. The changes in bird communities as you go up a tropical mountain are largely due to climatic differences, and the resulting changes in habitat that result from them. This is generally expected and was true in our study as well. Our main contribution, however, was to find evidence of diffuse competition (between groups of similar species) in ‘fine-tuning’ the ranges of individual species. This was particularly noticeable for bird species that live near the base of the mountain – they lived further up on mountains where fewer potential competitors were present on the upper slopes. This effect was less important in setting the lower range limits for birds living near the mountain tops, indicating that these mountain species may be stronger competitors than the lowland species (although others have found the opposite – we need to keep investigating the nuances of these interactions).
Next steps. Recent advances in Bayesian joint species distribution models now make it possible to combine many sources of information about these birds in a single modeling framework – traits, phylogenetic relationships, and environmental characteristics. My collaborators and I are in the process of combining our datasets of bird surveys across the island of Borneo with trait measurements from museums and forest information from remote sensing to better understand how the environment and species interactions shape bird communities.
Anything else? A description of the process, motivation, and rewards of this research wouldn’t be complete without a mention of the many local Indonesians and Malaysians who guided me through the forest and taught me so much. They have such a wealth of knowledge from a life spent in the forest and are a true highlight of time spent in Borneo.
Regarding myself, I’ll just say that my interests span beyond Borneo to a wide variety of questions in community ecology. Some other current projects include examining the effects of forest management practices on wood-living beetles in Scandinavia, and analyzing long-term time series of birds and moths from Africa, South America, and Europe.
If you could study any organism on Earth, what would it be and why? As an ecologist, I prefer to answer with a system rather than a single organism. Different taxonomic groups, whether birds, insects, plants, are often studied in relative isolation for practical reasons – just researching a single species is difficult enough! And yet we know that nature knows no such boundaries. Think of a tropical forest – fruit-eating bats, birds, and mammals are all relying on the same resources, and the lives of predatory insects and insectivorous bird and bats are closely linked as well (to give a very simplistic example). My ideal research program then would be to examine the interactions among plants, insects, birds, and mammals in the tropics – these areas are hyper-diverse, under-represented in the literature, and in immediate danger from rapid land use changes.