The Journal of Biogeography uses a structured abstract. Here are a few pointers on how to write one that will delight editors, reviewers, and readers.
Although less common that its counterpart the single paragraph prose-style abstract, the structured abstract has several benefits. One is that it helps readers find the key information that describes a study. Another is that it makes writing the abstract much more straightforward, guiding authors to include that key information, to align aims with methods and results and conclusions, and to avoid repetition (e.g. the location and taxon need only be mentioned once unless you are setting up contrasts among multiple locations/taxa).
As described by the journal’s Information for Authors, the structured abstract “should be no more than 300 words, presented as a series of factual statements under the following headings: Aim, Location, Taxon, Methods, Results and Main conclusions” (emphasis added). With the format set by these essential criteria, there is then some flexibility to provide different amounts of detail in each of the sections. However, the best structured abstracts also achieve a certain ‘balance’ and ‘parallel’ structure among sections, taking the reader from the context for the study, through what was done and found, to how this has advanced the field of research.
Thus, the journal’s Information for Authors goes on to note that the structured abstract’s Aim “should give a clear statement of the principal research question(s) or hypotheses,” that the Methods “should give details of materials/sampling/methods of analysis,” and the Main conclusions “should give the main take-home message.” Note that all of these things will benefit from being provided appropriate context, which we strongly recommend be used to set the scene for your study, i.e. as the first sentence of your Aim.
Although not stated explicitly, the Results section of the structured abstract should naturally bridge from the Methods — by reporting the outcomes of the analyses that justify the principal interpretations that lead — to the Main conclusions.
The journal’s Information for Authors also goes without saying that the Main conclusions should, of course, address the statements made in the Aim.
To better help you achieve these goals, to write a structured abstract that is maximally informative, we have provided something of a template below to act as a more explicit guide.
——————
The structured abstract (length guides are approximate, and must total ≤300 words):
Aim: A contextual statement explaining (1) the conceptual/theoretical context for the study, (2) the major knowledge gap to be filled, and (3) the specific aim of the study. The specific aim may be the principal research question(s) or hypothesis/hypotheses. (Approximate length: two to three sentences [40–50ish words].)
Location: The name of the area, place, region. (Approximate length: one or few words)
Taxon: Indicate the main group. (Approximate length: one or a few words.)
Methods: Give major details of materials, sampling, and methods of analysis that will communicate your study design and its robustness, especially with respect to answering the question(s), or testing the hypothesis/hypotheses, posed in the Aim. (Approximate length: several sentences [60–100ish words].)
Results: Give major details of the results of your analyses, especially those pertinent to your question(s) and hypothesis/hypotheses. There should be no inference nor interpretation (Approximate length: several sentences [60–100ish words].)
Main conclusions: State whether hypotheses posed in the Aim were refuted or supported; answer any questions asked. Interpret your results in the context of any questions posed. Address their implications for the field of study. (Approximate length: several sentences [60–100ish words].)
How efforts to understand diversity in urban areas could change perceptions of biodiversity in cities and increase green spaces.
Studies about community assembly typically focus on local community properties along natural-rural-urban gradients and neglect regional processes. As such, it remains unclear which functional traits are filtered when going from a regional pool of potential colonists to local communities in urban habitats. Also, urban community assembly studies often focus on one or few urban habitats whereas a city is composed of a mosaic of different habitats including urban parks, gardens, brownfields, and densely urbanized habitats. Tackling these questions is difficult because it requires detailed occurrence data about several hundreds of species occurring in a region and their functional traits. By assembling data from several studies in the city of Zurich (Switzerland), national scale occurrence data of bees and carabid beetles, and several trait databases we were able to study in great detail the regional processes underlying community assembly in the city of Zurich. The studies in the city of Zurich were carried out in four different urban green types (parks, domestic gardens, allotment gardens, green roofs, and brownfields) and used standardized sampling methods (pitfall traps and colored pans).
Image: Allotment gardens under pressure in the city of Zurich, Switzerland (Photo by David Frey)
———————— COVER STORY 47(3): Fournier, B, Frey, D, Moretti, M. 2020. The origin of urban communities: From the regional species pool to community assemblages in city. J Biogeogr. 47(3):615-629. https://doi.org/10.1111/jbi.13772 ————————
Cities are very particular settings for community assembly as they are highly heterogeneous and include a diverse array of small habitats occurring close to each other. Cities are thus likely to impose specific constrained on species niches and dispersal. Understanding these constraints is important because more and more people live in urban environments worldwide. In addition, the climate in cities is several degrees warmer and dryer than in natural habitats. As such, understanding the regional processes of urban community assembly can help create a better environment for human populations and remedy the negative effects of climate change.
During this research, we were surprised by the extraordinary species and functional traits diversity occurring in the city and the important differences in community assembly processes among taxonomic groups and urban habitats. Most importantly, our study revealed that species occurring in a broad range of natural habitats could potentially colonize the city. Only specialist species from alpine habitats had very low chances of successful colonization. Also, we did not find neophyte or invasive species despite the high chances of human-facilitated dispersal in cities.
An important question arising after the completion of this work is how different are regional urban community assembly processes from those occurring in more natural environments. Cities are generally thought to have a negative impact on biodiversity. However, we are wondering how the filtering of species and traits differ between urban habitats and more natural environments. Answering this question can help change people’s perception of biodiversity in urban settings and encourage people to increase the proportion of green habitats in cities worldwide.
Written by: Bertrand Fournier (1), David Frey (2), & Marco Moretti (3) (1) Concordia University; Department of Biology; Montreal, Canada. (2) Swiss Federal Research Institute WSL, Biodiversity and Conservation Biology; Department of Environmental Systems Science, ETH Zurich. (3) Swiss Federal Research Institute WSL, Biodiversity and Conservation Biology.
Emma Jardine is currently an environmental land management scientist at DEFRA. She is interested in the use of functional traits to understand how plants respond to the environment. Her recently published work in the Journal of Biogeogrphy considers the relationship between functional traits in grass species and environmental variables at a macroecological scale. Emma shares how the rich collection of plant specimens in Kew Royal Botanical Garden’s herbarium made her research possible.
(left) Emma Jardine, finding some time to visit the Great Plains and Rocky Mountains on a trip to think about grassland ecosystems and how they respond to drought at a DroughtNet data synthesis meeting, Fort Collins, Colorado. (middle) Herbarium specimen of Digitaria eriantha Steud. collected in 1927 in South Africa which is part of the collection at the Royal Botanical Gardens Kew. (right) One of the many rooms which form the herbarium at Kew botanical Gardens. The collection houses approximately 7 million herbarium specimens in total which have been collected from around the world over the past 170 years.
Institute. The featured research was conducted at the University of Sheffield.
Current role. Former postdoc now working on the environmental land management team at DEFRA.
Major research themes. Using functional traits to explain plant response to environment.
Current study system. I use grasslands as a study system to explore trait environment relationships. Grassy biomes are perfect for macro-ecological research as they cover between 20-40% of the terrestrial globe, have evolved to tolerate a wide variety of climatic conditions and soil regimes and have fewer dominant species than for example tropical forests. Grasslands play an important role in carbon sequestration, silica cycling and food security. These are important ecosystems but are threatened by land-use and climate change.
Recent paper in Journal of Biogeography.Jardine, Emma & Thomas, Gavin & Forrestel, Elisabeth & Lehmann, Caroline & Osborne, Colin. (2020). The global distribution of grass functional traits within grassy biomes. Journal of Biogeography, 47(3):553–565. 10.1111/jbi.13764.
Motivation of this paper. A central goal of ecology is to find rules to describe patterns in nature and how these patterns arise. The identification of patterns that determine vegetation dynamics at the global scale has the potential to improve our understanding of how ecosystems will respond to global change. Studies of global change often divide plants into functional types to make predictions without requiring detailed knowledge of species, with grasses being grouped by photosynthetic type (C3/C4). However, this approach ignores functional variation within these groups. We therefore wanted to know if relationships between functional traits and resource availability that are well known at community scale, could be applied over global scales.
Key methodologies. One problem that needs to be overcome when working at macro scale is sampling large numbers of globally distributed species. Trait databases are often biased towards woody taxa, so this makes accessing a large set of grass species grown in their natural environment across a global distribution very challenging. It would be a dream come true to travel to all the savannas, steppes, prairies and pampas of the world to assemble such a dataset. Unfortunately, this was beyond the timescale and budget of this research project. I instead developed methods for measuring functional traits from herbarium specimens and went on a worldwide tour of grassy biomes from within the herbarium at the Royal Botanical Gardens, Kew. I was amazed at the scale of the collection and there I found hundreds of specimens collected by intrepid explorers from surprisingly remote parts of the globe.
Unexpected challenges. Functional traits are usually measured from fresh material, so the first part of the project was to develop methods for taking measurements from dried material. Grasses can be notoriously tricky to identify, but one of the great things about using the herbarium (as opposed to field collection for this piece of research) was that the expert taxonomists at Kew had already done this job for me. One of the daunting aspects of this work was the number of specimens required to obtain a globally representative sample. Several weeks were spent just searching through the many thousands of specimens in the cupboards at Kew for the globally dominant species I was interested in, before any measurements for the final dataset could begin.
Major contributions. We conducted a comparative analysis of functional traits, climate and soil fertility for almost one third of the globally dominant grass species from all continents including representatives from all major grass phylogenetic clades. Global scale studies of trait environment relationships commonly span broad taxa whereas ours was specific to grasses. Previous studies have also not accounted for the influence of phylogeny on observed patterns. We found that relationship between traits and environment are weak. This is because trait variation is greater within than between different vegetation types. We also discovered a strong phylogenetic signal in all of the leaf traits measured. We therefore concluded that evolutionary history provides a stronger explanation of trait distribution than contemporary environment.
What is the next step? We found it interesting that traits varied more within than between different vegetation types and this result led naturally to research focused on processes that vary over smaller scales than climate, for example fire and herbivory. In collaboration with other colleagues we have been using this dataset to contribute to work investigating how globally fire filters functional traits. We have also been looking at the relationship between traits, fire and mammals across Africa. Through this we hope to gain more of an understanding of the processes that are responsible for structuring grassy ecosystems, which will in turn, provide information on how to conserve them.
If you could study any organism on Earth, what would it be and why? Grasses of course! I’m fascinated by Savanna ecosystems as they are prone to so many disturbances, for example fire, drought and herbivory. I’m particularly interested in the effects of herbivory on vegetation dynamics. As the only continent with an extant megafauna African savanna are the ideal system for studying this and provide the perfect excuse to research both plants and animals.
Kristen de Graauw is a dendroarchaeologist: that’s someone who studies the plant material in old human-made structures, such as buildings, artefacts and furniture. She is a recent PhD graduate from West Virginia University. In her recent work in the Journal of Biogeogrpahy, she describes the role of human occupation in shaping the distribution and dynamics of North American forests. Kristen shares how these insights were made possible from analysis of tree rings in historical log buildings.
Kristen K. de Graauw climbing into the hayloft of a late 1700s barn to inspect logs for sample collection.
Institute. Recent graduate from West Virginia University.
Academic life stage. PhD (graduated 2019).
Major research interests. Dendroarchaeology, Forest History and Ecology, Physical Geography, Environmental Science.
Current study system. Currently, my major research focus is on using tree-ring data from historic log buildings (typically Quercus alba, Liriodendron tulipifera, Pinus species) to explore forest history in eastern North America. This is a relatively new area of research exploration, and I’m finding that historic log buildings hold great potential for recording forest dynamics during the 1500s-1800s, a period not well represented in live tree records in eastern North America. My specific interests are related to changes in land management from Indigenous to European immigrant occupation and how those changes shaped our understanding of modern forests.
Motivation behind this paper. My co-author and I were compelled to undertake this project after reading Koch et al. (2019) in Quaternary Science Reviews. In this study, the authors hypothesized that there was mass forest regrowth in the Americas following the depopulation of Indigenous Peoples and that the increase in biomass ultimately led to the coldest portion of the Little Ice Age. A fascinating study! I was already using tree-ring data from historic log buildings to reconstruct forest age structure and disturbances during the European immigration period and recognized the potential of using my data to address the forest regrowth portion of the Koch et al. (2019) hypothesis. European immigrants’ historic log buildings were constructed in areas that were likely previously inhabited by Indigenous Peoples, so they were an ideal data source to use in this investigation of potential land management change.
(left) The Barracks, a late 1700s log building used in this study. (right) The Pitsenbarger farm, an example of historic log homesteads from the mid-1800s in the central Appalachian Mountains.
Key methodologies. Until recently, tree-ring data from historic log buildings were not generally used to investigate ecological questions. My dissertation provides one of the first formal introductions to the topic and this paper provides the first application (I believe) of historic logs to address an ecological question. We collected samples from 18 log buildings and compared recruitment dates (estimate of tree establishment), early radial growth (fast versus slow growth in first 100 years), and growth release events (an indication of canopy status, i.e. “open” or “closed”) with 8 old-growth forests to test for evidence of reforestation. We hypothesized that if there was evidence of forest regrowth following depopulation it would be present in historic log buildings because they represent areas suitable to human occupation, but not present in old-growth forests because these sites are generally located in areas not suitable for habitation.
Unexpected challenges. One of our greatest challenges was locating the specific tribes that inhabited the Ridge and Valley region of the central Appalachian Mountains in the 1600s. We wanted to accurately present the tribes that lived in this region at that time, and more specifically we wanted to understand their unique land management strategies. However, published records of the Indigenous Peoples at that time are relatively sparse and we were left with broad language groups that likely inhabited the region. We have since been informed about the Native Land App (native-land.ca), which provides a user-friendly map interface for locating territory boundaries of tribes in North America. While there are still some gaps in the data in our region, we now have a better understanding of who lived here in the 1600s and can begin to explore their land management techniques with the help of archaeologists and ethnographers.
Major results and contributions to the field. By comparing initiation, growth, and release events of historic logs (representing habitable areas) versus old-growth forests (representing areas unsuitable for human habitation), our data demonstrates that there were significant differences in tree growth in these two site types and that the timing of tree initiation coincided with the estimated timing of Indigenous depopulation from the region. However, tree recruitment dates across site types (historic buildings and old-growth forests) were also consistent with a sub-continental drought event in eastern North America, which may have led to tree mortality and subsequent recruitment. If we can replicate these results across a broader area in eastern North America, specifically in places where the sub-continental drought did not occur and/or where depopulation occurred earlier or later than the drought, it would suggest that forests encountered by European immigrants were largely created through cessation of large-scale land management at the time of European contact and depopulation (migration, mortality) of Indigenous Peoples.
The next steps in this research? The next step is to repeat our study using tree-ring data from historic log buildings in other portions of eastern North America. We are specifically interested in working in the southeast and along the coastal region. In doing this, we will test for forest regrowth and compare it with the estimated timing of depopulation in these regions, which would be different from the timing of depopulation in the central Appalachian Mountains. If we find evidence of forest regrowth that is coincident with estimated depopulation in the southeast and in coastal regions, it would suggest that Indigenous depopulation led to major forest changes in eastern North America. Such a finding would also contribute evidence to the “cooling” hypothesis posed by Koch et al. (2019) if evidence of reforestation is contemporaneous with the nadir of carbon drawdown (circa 1610 CE).
(left) Sanded white oak (Quercus alba) cores collected from a historic log building in the central Appalachian Mountains. (right) Kristen K. de Graauw collecting core samples from logs in a barn.
If you could study any organism on Earth, what would it be and why? Well, trees are my “thing”. I love mighty oaks, specifically white oak (Quercus alba), so I already study my favorite organism! But it would be amazing to work with older wooden archaeological materials/artifacts in other parts of the world.
Any other little gems you would like to share? My colleagues like to give me grief about how easy it must be to collect samples in a log building versus a forest, but what most people don’t consider is the massive amount of wildlife (spiders, snakes, mice, raccoons, etc), dead wildlife (see wildlife – and a cow!), animal feces, rusty objects, and general accumulation of human artifacts over centuries that I encounter in these historic buildings. Each building has its own theme, or “personality”, and some are more horrendous (and/or hazardous) than others. The most challenging building I worked in, though, was a museum full of beautiful antiques. I worry enough about carefully collecting cores from these centuries-old logs – the last thing I want to worry about is getting sawdust on a priceless quilt!
Esther Dale studies plant diversification as a postdoc at Manaaki Whenua – Landcare Research in Dunedin and the University of Otago, Department of Botany. Her recent publication in the Journal of Biogeography tests biome conservatism in Australian Acacia using species distribution modeling. Esther discusses the implications of her findings, particularly that hyper-diverse Australian Acacia provides a rare example of a plant lineage in which most species occur across multiple biomes.
Esther Dale in the Palm House at the University of Copenhagen Botanical Garden
Institution: Manaaki Whenua – Landcare Research, Dunedin; University of Otago, Department of Botany
Current academic life stage: Postdoc
Research interests: I study plant diversification, focusing on the role of shifts between different biomes in lineage evolution.
Current study system: Australian Acacia—they’re hyper-diverse, with over 1000 species, making them one of the most diverse vascular plant genera globally. Australian Acacia have managed to occupy many different, contrasting ecosystems such as desert and tropical rainforest.
Recent paper in Journal of Biogeography: Dale EE, Larcombe MJ, Lee WG, Higgins, SI (2020). Diversification is decoupled from biome fidelity: Acacia–a case study. Journal of Biogeography, 47(2):538– 552. DOI: 10.1111/jbi.13768
Motivation for the paper: We were interested in examining how plant lineages evolve in relation to the biomes that they occupy. Biome conservatism, the tendency for lineages to remain in their ancestral biome, is regarded as relatively typical in plant lineages. We wanted to explore whether biome conservatism constrains diversification by keeping lineages within a particular biome, if diversification is primarily occurring within biome boundaries, and if diversification is associated with specialisation to a single biome. We decided to use Australian Acacia to test this because it is hyper-diverse (over 1000 species!), it has a published phylogeny, and distribution data were available.
Acacia aneura in Mallee. This vegetation would be classified as Mediterranean (WWF Biomes), Short Low-productivity Non-seasonal (Functional Biomes), or Eremaean (Crisp Biomes and González-Orozco Biomes). Photo credit: Matthew Larcombe.
Key methodologies: We used species distribution modelling to identify which biomes each species occurs in. We had noticed with the distributional data that some biomes were under-sampled, which might make species seem more specialised to certain biomes than they are in reality. To minimise this source of bias, we used predicted distributions rather than distribution records to determine the biomes occupied. In addition, there are a variety of different biome maps with different numbers of biomes. Previous biome conservatism work has used a variety of different biome maps and we thought it was likely that the biome typology used would influence the conclusions being drawn. We used four different biome typologies to check that our findings were robust over multiple different biome concepts.
(left) Esther at the Brisbane Botanic Gardens Mt Coot-tha examining Acacia disparrima. (right) Esther at Kings Plains National Park, New South Wales. The vegetation in this area would be classified as Temperate Forest (WWF Biomes), Tall High-productivity Non-seasonal (Functional Biomes), Southeastern Temperate (Crisp Biomes), or Euronotian (González-Orozco Biomes). Photo credit: Zoë Stone.
Unexpected challenges: In contrast to the expectation under biome conservatism, we observed that most species (91%) occurred in multiple biomes. This was quite surprising because it contrasts some previous work demonstrating biome conservatism as widespread. It indicates that specialisation of species to a single biome cannot be assumed, and analyses examining biome shifts and biome conservatism should allow for species that occupy multiple biomes. The main challenge with this research was the size of the dataset. It involved 481 species, 151735 occurrence records, and global environmental data layers with a 1 km resolution, meaning we needed to be efficient with our analyses. Most of us working on this project hadn’t worked on Australian species before and were not familiar with the history of the flora or climate, so it was challenging taking on such a characteristically Australian group without much experience of Australian ecosystems. However, it was probably also useful—we hope—to be able to contribute a fresh perspective.
Major result and contribution to the field: We found a consistent pattern of cross-biome diversification with all four biome typologies. Higher diversity clades had greater niche size, indicating that diversification is linked to occupying new niche space rather than partitioning the ancestral niche. Our work demonstrates that Acacia can easily overcome biome boundaries, and diversification in Acacia is not constrained by biome conservatism. This contributes a rare example of a lineage where most species occur in many biomes, with diversification occurring across biome boundaries. Our findings also indicate that lineages can have many species that occur in multiple biomes, so analyses should allow for species that occupy multiple biomes.
What are the next steps? We are interested in applying what we’ve learned from looking at Acacia, with its large spatial and taxonomic scale, to New Zealand plant lineages. We are keen to follow on from these findings by exploring how diversification is affected by new biomes appearing, and whether biome shifts are associated with trait innovations. New Zealand will provide an interesting contrast to our Acacia work because there are fewer biomes and smaller focal lineages. In New Zealand there has been a clear sequence of different biomes becoming available, which presents an excellent opportunity for examining the role of new biomes and biome shifts in lineage evolution. New Zealand is a fantastic system for testing hypotheses about plant evolution because of the quirks of the flora, like high incidences of polyploidy, divarication, and small inconspicuous flowers, and its isolation, making it a natural experiment in evolution.
If you could study any organism on Earth, what would it be and why? I would love to be able to study subantarctic megaherbs. The subantarctic islands are known for their megaherbs, which tend to be much larger and have flowers that are brighter in colour than their New Zealand mainland relatives. It would be fun to examine their evolutionary history to understand how they have adapted to the conditions in the subantarctics, and what selection pressures are driving these changes.
Any other little gems you would like to share? This type of work would not be possible without the excellent body of herbarium specimens in Australia, so a big thanks to all the collectors, herbarium curators, and the Atlas of Living Australia who do such important work!
Eucalyptus regnans forest in Gippsland Victoria. This vegetation would be classified as Temperate forest (WWF Biomes), Tall High-productivity Non-seasonal (Functional Biomes), Southeastern Temperate (Crisp Biomes), or Euronotian (González-Orozco Biomes). Photo credit: Matthew Larcombe.
Writing the perspective Why Mountains Matter for Biodiversity (Perrigo et al. 2020) was a chance for myself, along with Carina Hoorn and Alexandre Antonelli, to explore and distill some of the ideas that came up while editing a book that was published two years ago: Mountains, Climate and Biodiversity (Hoorn et al., 2018).
One of many early versions of a figure from the paper that were exchanged by the authors to make sure all the processes they showed were as accurate as possible.
Mountains are increasingly discussed in relation to biodiversity and, more specifically, biogeography. This is logical: mountains are much less affected by direct human activity as compared with lowlands, meaning they tend to be more “natural” systems to study. They are important when it comes to climate change as well. Often, a population must move a shorter distance to track a suitable habitat on a mountain, as the climatic variation from increasing altitude parallels the changes seen in increasing latitude in many ways.
———————— EDITORS’ CHOICE 47(2): Perrigo, A, Hoorn, C, Antonelli, A. (2020) Why mountains matter for biodiversity. Journal of Biogeography 47(2): 315–325. https://doi.org/10.1111/jbi.13731. OPEN ACCESS ————————
Critically, in the coming decades mountains will play a key role in biodiversity conservation as refugia for many species. These “Anthropocene nunataks” will be surrounded by seas of land that is exploited by humans, making a barrier to dispersal that may be analogous to the ice sheets that surrounded ice age nunataks.
While hiking up Mt Bisoke in Rwanda (ca. 3700 m), it is possible to observe a progression of vegetation types—influenced by the changing altitude—in a matter of hours.
In our perspective we outline what we see as some of the biggest challenges we now face in moving the study of mountain biodiversity forward. Among these are the relatively low availability of sufficiently specific data compared to lowlands on, for example, climate, soil-type and vegetation patterns. We also express the need for more studies that incorporate both theoretical and empirical aspects to push method development forward, while highlighting good examples.
Trans- and inter-disciplinary projects are encouraged throughout academia to explore novel perspectives. But often these are a challenge in practice because we are “speaking different languages,” to steal a phrase I have heard repeatedly about this type of work. Even though geology and biology are just a stone’s throw from one another, there are mountains between us (pun intended…). This became increasingly apparent while working with the various chapters of the book, where differences in terminology and tradition often led to misunderstandings. These were frequently a result of mutual knowledge gaps.
The table in our perspective is one of our most concrete efforts to overcome this. It summarizes the different methods used for dating mountains. The concept is simple: different methods are used to figure out just how old mountains are. But in practice, especially for biologists, it can be daunting to understand these methods in context and relative to one another. What ages are they good for? When would you use one or the other? Where can you even look to figure out what they mean or how they work, in basic terms?
However, in the beginning this information wasn’t laid out as a table. Carina Hoorn was the mastermind behind the mountain dating overview, but it was initially a long text with a repeating pattern: dating method, how it works, when it is best used, ages it is relevant for, references, and so on. It was extremely informative, but hard to navigate. After some discussion we realized: maybe this information would be better presented as a table? In the end, it was much easier to both read and interpret this way. The mountain dating table is a cornerstone of this perspective: we hope this accessible overview opens up doors for further cross-disciplinary understanding.
We see a value in reflecting on a project after its completion, especially one with as many contributors as Mountains, Climate and Biodiversity. Like Humboldt, whose work was among our constant inspirations for this perspective, we attempted to find key themes in a massive amount of information from various disciplines and from this take the parts we found the most interesting, useful and provocative.
Finally, we are encouraged by the growing interest in mountain biodiversity we have seen over the last few years. We have received positive feedback and started new discussions stemming from both Mountains, Climate and Biodiversityand the early online version of our perspective. This issue of the Journal of Biogeography features three other mountain-themed papers (Brambach et al. 2020, Li et al. 2020 and Maicher et al. 2020), and follows on the heels of the 2019 special issue celebrating Humboldt’s 250-year birthday and the legacy of his work.
Written by: Allison Perrigo.
Director, Gothenburg Global Biodiversity Centre, University of Gothenburg, Gothenburg, Sweden. @DrSlimeMold, @GGBC_GU
I would like to thank Carina Hoorn (@carinahoorn), Harith Farooq, and Ferran Sayol who all provided feedback on this blog post, as well as Alexandre Antonelli (@antonelli_lab) for discussions on the content. All three co-authors (AP, CH, AA) are grateful to all of the authors who contributed to Mountains, Climate and Biodiversity
Friederike Gebert is a postdoc working in tropical entomology at the University of Würzberg. Her recent work, published in the Journal of Biogeography, focuses on understanding the patterns and drivers of dung beetle distribution along an elevational gradient on Mt. Kilimanjaro. From the collection of dung beetles to the measurement of mammal dung resources, Friederike provides an insider look into the study of two very different but functionally related groups of organisms on the highest free-standing mountain in the world.
Friederike whilst setting up a pitfall trap at the base of Mt. Kilimanjaro at 870m asl on a study plot located in the savannah
Institution: University of Würzburg, Department of Animal Ecology and Tropical Biology
Current academic life stage: Postdoc
Research interests: I am interested in tropical entomology and in the impact of elevation and land use on species’ distributions, traits, and ecosystem services.
Current study system: Currently, I am studying dung beetles and mammals on Mt. Kilimanjaro, Tanzania. The fascinating thing about my research is that I combine the study of two very different, yet closely functionally related groups of organisms on the highest free-standing mountain in the world to investigate the drivers of biodiversity along elevational gradients. Both dung beetles and mammals are of immense importance in ecosystems, the former because they provide many ecosystem services related to dung decomposition, and the latter because they turn over high amounts of biomass and provide an array of services from pollination to the maintenance of habitat heterogeneity.
Recent paper in Journal of Biogeography: Gebert F, Steffan-Dewenter I, Moretto P, Peters MK. (2020) Climate rather than dung resources predict dung beetle abundance and diversity along elevational and land use gradients on Mt. Kilimanjaro. Journal of Biogeography, 47(2):371–381. https://doi.org/10.1111/jbi.13710
Motivation for the paper: The distribution of species along mountains has fascinated biologists for more than two centuries. Even though the patterns of species diversity along elevational gradients are well described, the drivers behind them remain controversial until today. Many hypotheses are discussed, a general disagreement being whether energy-related or climate-related drivers are more important. I wanted to contribute to this current discourse on the predictors of biodiversity by investigating the patterns and drivers of ectothermic organisms—dung beetles—along a huge elevational gradient on Mt. Kilimanjaro. One shortcoming of studies investigating the importance of energy versus the importance of temperature in driving biodiversity gradients is that often, energy resources are not directly measured. Instead, primary productivity is used as a proxy for resources. However, most organisms use specific food resources which may not be represented accurately by primary productivity. Therefore, we tried to measure the energy resources available for dung beetles in the field.
(A)
(B)
(C)
(A) The study system: Mt. Kilimanjaro – this photograph was taken at 1600m from the research station, Nkweseko, which served as the base for the dung beetle study. It shows the south face of Kibo, the summit of Mt. Kilimanjaro at 5895m. Nkweseko research station is located in the cultivated zone of Mt. Kilimanjaro inside the coffee-banana belt. The predominant form of cultivation here is traditional agroforestry of the Chagga people, which is called Chagga homegardens. (B)Here you can see very rich lower montane forest. This beautiful valley is located at 1600m and, unfortunately, is not part of Mt. Kilimanjaro National Park. Therefore, logging is commonplace and a threat to the preservation of this remarkable area. Here, two study plots were located.(C) Here you can see a camera trap set up in the forest. (below) A part of our collection. In total, we collected 10432 dung beetles belonging to 135 species.
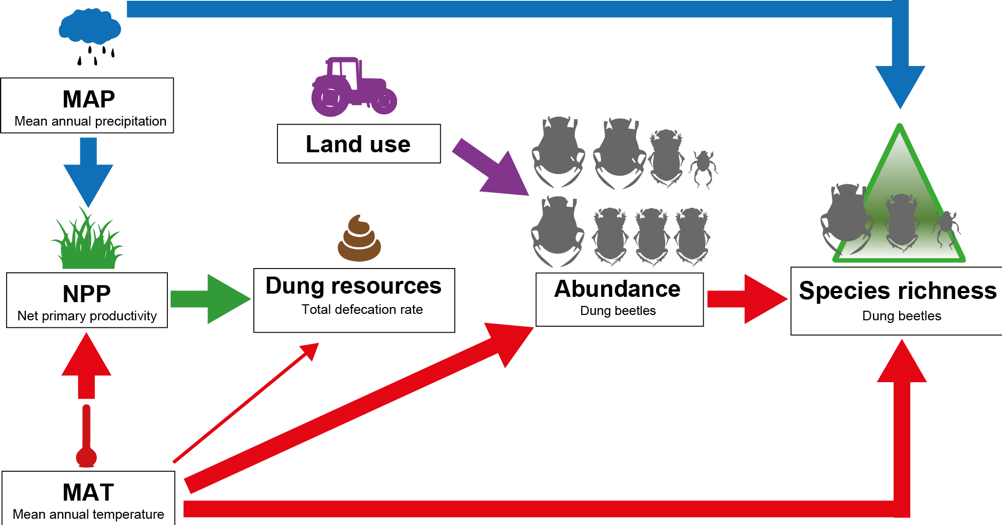
Key methodologies: We used baited pitfall traps to collect dung beetles. For measuring dung resources available for dung beetles, we calculated mammal defecation. To get the mammal data, we installed camera traps on the same 66 study sites along an elevational gradient of 3.6km on Mt. Kilimanjaro on which we collected dung beetles. We then estimated mammal defecation from mammal biomass data. To the best of our knowledge, this is the first study to accurately measure the amount of dung resources available for dung beetles. Apart from dung resources, we also investigated the impact of temperature, precipitation, land use and area on dung beetle species richness and abundance by means of path analysis.
A graph illustrating the drivers of dung beetle species richness and abundance. While dung beetle diversity is impacted by climate (temperature and precipitation), the main predictor for mammals (here represented as the dung resources) is net primary productivity, demonstrating the contrasting drivers of endothermic and ectothermic diversity.
Unexpected challenges: Since dung beetles rely upon mammalian dung as a food source for both adults and larvae and are thus closely functionally related to mammals, we expected mammalian dung resources to be one major driver of dung beetle diversity on Mt. Kilimanjaro. As we tried our best to have a good measure of dung resources for dung beetles, we were surprised to find that dung resources did not play a role in predicting dung beetle diversity. Since most dung beetles are active during the rainy season, I mainly collected dung beetles during the period of the long rains on Mt. Kilimanjaro. Because of muddy roads and paths, it was not always easy to reach the study plots and we had to overcome challenges like trees blocking the road or cars stuck in the mud.
Major result and contribution to the field: We found that even though we tried to meticulously measure the actual dung resources available for dung beetles, temperature was the major driver of dung beetle diversity on Mt. Kilimanjaro. This study shows that along huge environmental gradients, temperature is the main driver of diversity for ectothermic organisms. However, when we looked at the drivers of mammals, which are endothermic, we found that energy resources were the most important drivers. To conclude, our study is an important contribution to the current debate on the drivers of biodiversity as it illustrates that the drivers for montane diversity depend on the thermoregulatory strategy of organisms. Furthermore, our finding that dung beetle diversity is driven by temperature has important implications for biodiversity conservation in the context of climate change, as rising temperatures may have negative repercussions on dung beetles and their associated ecosystem services.
What are the next steps? The next step in this research is to look at the ecosystem services provided by dung beetles to investigate whether climate or biodiversity is more important in shaping ecosystem services along environmental gradients.
If you could study any organism on Earth, what would it be and why? In ecology, we are always in search of general patterns. I especially like organisms that make us question the concept of generality and make us acknowledge that there are always exceptions to the rule, which is one of the most intriguing features of studying biodiversity. That is why, even though I am actually an insect person, I would love to study the Grasshopper Mouse (Onychomys sp.). This mouse is so special because, in contrast to other rodents, it relies on a carnivorous diet and even feeds on scorpions and snakes, being immune to their venom. And since I am an old Dracula fan, I love the fact that this mouse is nocturnal and howls to the moon like a proper werewolf mouse!
Any other little gems you would like to share? During my research on Mt. Kilimanjaro, we collected a total of 80,000 film snippets from camera traps. Whilst most of the videos showed moving grass in the savannah, around 1600 videos did actually show mammals. Our biggest mammal highlight was that we filmed the elusive antelope species Abbott’s Duiker (Cephalophus spadix) for the first time on Mt. Kilimanjaro. This antelope is endemic to Tanzania and only occurs on few isolated mountain massifs. Up to now, the distribution of this species on Mt. Kilimanjaro has not been known – now our research suggests that Mt. Kilimanjaro might be a stronghold for this species. What’s cool is that we were the first ever to film an Abbott’s Duiker pair and a male trying to mate.
(left) Alpine Helichrysum scrub vegetation at 3800m. (right) The highest study plots were located at 4500 m.
How efforts to understand origins and diversity led to efforts to conserve and protect the world’s largest bats
Emergence of Pteropus vampyrus from a colony on Flores, Indonesia
———————— COVER STORY 47(2): Tsang, SM, Wiantoro, S, Veluz, MJ, et al. (2020) Dispersal out of Wallacea spurs diversification of Pteropus flying foxes, the world’s largest bats (Mammalia: Chiroptera). J. Biogeography 47(2): 527– 537. https://doi.org/10.1111/jbi.13750 ————————
Long-distance dispersal (LDD) is often regarded as a rare and unpredictable event, causing some to consider it an untestable hypothesis. However, advances in genomics, niche modeling, and computation enable direct and indirect testing of LDD hypotheses, particularly in understanding the evolution of recently diverged taxa. These studies have primarily been conducted on plants and a handful on bird groups, but the role of dispersal in bat diversification is scarcely studied. Historically, research on Paleotropical bats has lagged behind other vertebrates, but recent capacity-building efforts have enabled internationally collaborative research in Southeast Asia. Studying LDD in flying foxes, the world’s largest bats in the genera Pteropus and Acerodon, is difficult because some widespread species are found in more than half a dozen countries, while numerous endemic species are restricted to individual islands or island groups from American Samoa to Zanzibar. By assembling a multi-national team for locating and sampling tissues from these rare bats, we were able to conduct one of the first biogeographic studies of a species-rich mammal taxon in the Indo-Australian Archipelago (IAA).
The IAA is a fascinating biogeographic laboratory, as the fission and fusion of landmasses over geologic time have both isolated and re-connected areas in rapid succession. Biotic studies in the IAA are few relative to that of other tropical areas, particularly in Wallacea, the area between Borneo and New Guinea. Many islands in Wallacea have never been connected to a continent. Our study suggests that Pteropus originated in Wallaceaat a time when the relatively depauperate ecosystems on recently formed islands presented “blank canvases” to which flying foxes dispersed and subsequently diverged from their ancestors. For the most part, the number of flying fox species found in an area seems unaffected by the size of the bat, the distance between islands, or the age of the lineage (with a possible exception in the South Pacific). Flying foxes can carry fruits, swallow seeds whole, and fly nearly 90 km in an evening, potentially over water. They are acknowledged as crucial seed dispersers in the rainforest where they occur, but their impact on plant historical biogeography and diversification, through co-dispersal, for example, is little studied.
During the course of fieldwork for this study, members of our team located bat species that hadn’t been seen alive in over two decades. We also found that colony sizes are shrinking throughout the IAA; a colony’s crepuscular emergence no longer blankets the sky with black, leathery wings. We observed that species throughout Asia are hunted at unsustainable levels for human consumption, which creates a conservation crisis for bats and a public health hazard for humans, who could be exposed to potentially lethal pathogens. This has motivated us to train multiple grass-roots organizations and local scientists in standardized population monitoring and outreach methods, along with push for localized hunting bans or regulations to prevent further decline. Expansive biogeographic studies like this may not be possible in the future if the current rate of population extirpation continues. Flying foxes and other fruit bats are charismatic but little known. They are perhaps best studied in Australia where roosts are near urban areas, but only 5 of the 65 known species are found there. A majority of species are endemic to single islands or island groups, and these species are understudied, despite their importance for forest regeneration on isolated, oceanic islands. Future studies will examine the environmental impact of flying foxes on terrestrial and marine ecosystems, as many species roost in large aggregates in liminal areas, such as mangroves, that abut vulnerable ecosystems such as coral reefs.
Written by: Susan M. Tsang (1) & David J. Lohman (2) (1) Research Associate, Department of Mammalogy, American Museum of Natural History; Research Associate, Mammalogy Section, Philippine National Museum of Natural History. (2) Associate Professor, Department of Biology, City College of New York & PhD Program in Biology, Graduate Center, City University of New York; Research Associate, Entomology Section, Philippine National Museum of Natural History (Philippines).
Rob Smith is a postdoc in the Department of Botany and Plant Pathology at Oregon State University, who studies the effects of changing atmospheric regimes on forest vegetation. Rob’s recently published work in the Journal of Biogeography discusses how epiphytic macrolichen vulnerability to climate change can signal atmospheric stresses among a group of organisms that many assume to be indifferent to climate.
Institution: Oregon State University, Dept of Botany and Plant Pathology
Current academic life stage: Postdoc
Research interests: Forecasting how global changes to atmospheric regimes alter the functioning, persistence and distribution of forest vegetation.
Current study system: Epiphytic lichens – perched upon the branches and trunks of other plants – are tightly linked to the atmosphere. Despite their mistaken “extremophile” reputation, these lichens live at the mercy of atmospheric nutrients, temperature and moisture. Climate, therefore, imposes strict limits on individual performance and geographic distributions. Since most lichens fit in the palm of the hand, they make excellent travel companions, in addition to being well-suited for reciprocal transplants and common garden experiments to better anticipate climate responses.
Motivation for the paper: We previously looked at climate-driven historical changes in lichen community compositions using a large-scale national forest inventory (FIA, the US Forest Service’s Forest Inventory and Analysis). Yet, we still lacked the basic ability to forecast future changes at large scales. Our new vulnerability approach arose from the observation that populations at the physiological “edges” of a species’ realized niche can also reveal its geographic edges – places on the landscape where incremental warming or drying could push a population to local extinction. For many species together, co-occurrence of such “vulnerable” populations would signal a community on the verge of compositional turnover. Forest workers would like to anticipate the location and magnitude of compositional changes before they occur.
(A)
(A) Climate stress from prolonged drying or warming can change lichen community compositions, including these pale green Alectoria hair-lichens draping western hemlocks in the mist near Cascade Pass, Washington, USA. (B)Bryoria horsehair lichens festoon a subalpine fir, with the glaciers of Mount Rainier in the distance, Washington, USA. Direct exposure to the atmosphere makes epiphytic lichens sensitive to climate variation. (C) Charismatic epiphytic lichens (like this “oakmoss” Evernia prunastri) are easily transplanted, making them ideal for examining climate tolerance. (D) Installing cameras for daily census of lichen dynamics as part of new work with the Epiphytic Lichen Observation Network (ELON).
(B)
(C)
(D)
Key methodologies: Ecologists often depict environmental responses as community-weighted means (e.g., temperature optima). Yet, central mean values ignore the “tails” of realized niches – boundaries which define the outer limits of persistence. By contrast, our vulnerability scores are explicit about how far populations are from their climatic edges, and therefore the conditions under which incremental climate changes would lead to local declines. Across 400+ epiphytic macrolichen species, and combined with climatic exposure at thousands of FIA plots nationwide, vulnerability scores let us identify geographic “hotspots” of expected compositional changes.
Unexpected challenges: One challenge was to adequately depict the realized climatic niche of several hundred species, including species whose ranges extended well beyond the study area. We resolved this by introducing hundreds of thousands of herbarium records from all of North America, from tropical to polar regions. This introduced its own challenge: how best to standardize the unequal sampling efforts typical of natural history collections? For this, we binned observations into spatial grid-cells of gradually increasing sizes, finally arriving at an acceptable cell-size that “smoothed away” unequal sampling efforts while preserving climate information.
Major result and contribution to the field: We found remarkably high climate-change vulnerability among a group of organisms that many folks assume are indifferent to climate! This supports the emerging perspective that epiphytic macrolichens can sufficiently signal atmospheric stresses. Unexpectedly, we also found that communities most vulnerable to warming were concentrated in low‐elevation and southerly locations. This suggests that warm-edge communities – commonly assumed to be “thermophilic” – may in truth be perilously close to exceeding their climatic limits. Could the vulnerability approach work for the organisms you study? You can try it for yourself, using the freely available R package: https://github.com/phytomosaic/vuln.
What are the next steps? We are working to generalize the vulnerability concept to admit multiple drivers that interact. This is because interactions among atmospheric stressors (climate and nitrogen excesses) and disturbances (wildfires) not only violate species’ tolerances directly, but can also aggravate pest/pathogen risks. We also need to compare vulnerability among different lineages and morphogroups (forest trees, shrubs, forbs, graminoids, lichens) to identify the most responsive organisms. Finally, we are developing user-friendly, open-source mapping tools that let forest workers quickly identify hotspots of compositional changes for adaptation purposes.
If you could study any organism on Earth, what would it be and why? To me, the most fascinating organisms are forest trees, which breathe our air and cycle our carbon. Even for these everyday organisms, there is some hint of mystery in simple acts like passive water uptake (solely by transpiration! no active pumping!). Despite being the dominant characters in forests, we are still far from accurately forecasting tree growth, survival and reproduction over large scales in the face of global changes.
Any other little gems you would like to share? It’s always fun to hear origin stories, how each person first began to engage the natural world. My first real entry to forest thinking was the simple result of being an introverted kid – stealing off down the leafy railroad tracks paralleling the Patapsco River to decipher Lao Tzu or Albert Schweitzer under red oaks. And then, looking up at red oaks, wondering. There are hundreds of ways to enter your own forest.
Vulnerability scores combine local climate exposure with sensitivities of individual species, like these Bryoria horsehair lichens above White River at Mount Rainier, Washington, USA.
Rafael Venegas is an ecologist with a passion for plants. He is currently a postdoc at the University of Alcalá. He uses phylogenetic methods to address questions in macroecology and biogeography to ultimately understand what shapes biodiversity and ecosystem services. In his recent paper with the Journal of Biogeography, he extends theory on phylogenetic community structure through specific consideration of phylogeny branching patterns. Rafael shares how insights on community assembly can be gained from this new analytical framework.
Rafael Molina Venegas. PhD and current postdoctoral researcher at the University of Alcalá (Madrid, Spain), posing in front of a pine forest (Pinus pinea) in the surroundings of Doñana National Park (southwestern Iberian Peninsula)
Research interests: Phylogenetics, macroecology, biogeography, plant biodiversity, ecosystem services.
Current study system: I’ve been working on Mt. Kilimanjaro flora for two years. Kilimanjaro is the highest single free-standing volcanic massif in the world, and includes lush jungles, cloud forests and cold and fire-adapted bushlands and scrublands that spread until the glacier’s domain at 4500 m. This montane vegetation stands in splendid isolation above the surrounding plains, where savanna woodlands and agricultural fields dominate the landscape. The geographical isolation of Kilimanjaro makes its people highly dependent on natural resources (the so-called ecosystem services), creating an interesting socio-ecological context that inspired me to design the project I’m currently leading. This project aims to explore connections between ethnobotanical knowledge (in my opinion one of the most palpable proofs of the reality of ecosystem services) and global plant biodiversity from an evolutionary perspective.
The truth is that phylogenies have always been in the background of my research agenda, including the development and refinement of phylogenetic methods for the study of biodiversity. Indeed, some of my ongoing collaborations concern spatial phylogenetics with a focus on the endemic flora of the Iberian Peninsula (~1975 species and subspecies, 27% of the vascular flora of the region), an outstanding plant biodiversity hotspot in the western Mediterranean. I am particularly interested in evaluating the role that soil conditions have played in the diversification and maintenance of this flora.
(left) Rafael, exploring the forests in the foothills of Mt Kilimanjaro. (right) A chamaephyte community studied by Rafael in the Iberian Peninsula, a hotspot for plant biodiversity in Europe. Amidst the Quercus oaks, purple lavenders (Lavandula pedunculata) and white-flowered gum rockroses (Cistus ladanifer) bloom in the Mediterranean sunshine.
Recent paper in Journal of Biogeography: Molina-Venegas, R., Fischer, M. & Hemp, A. (2019) Disentangling the fundamental branching patterns of phylogenetic divergence to refine eco‐phylogenetic analyses. Journal of Biogeography, 46, 2722-2734. https://doi.org/10.1111/jbi.13692
Motivation for the paper: My first steps in science took me to the field of eco-phylogenetics, which aims to infer community assembly mechanisms by means of the footprint they left on the phylogenetic structure of communities. For example, little phylogenetic divergence (i.e. clustering) may indicate community structure is shaped by environmental filtering, which is a major mechanism in harsh habitats such as Mediterranean saline soils, where closely-related salt-adapted lineages predominate (e.g. Tamaricaceae, Frankeniaceae, Amaranthaceae). However, classical indices of phylogenetic divergence disregard much of the biological information encoded in the phylogenies, because they are simply “blind” to the exact branching pattern of phylogenies. This is problematic because it precludes understanding of how ecological processes affect evolutionary relationships within communities. The prospect of overcoming this methodological shortcoming was the main motivation to work on this paper.
Key methodologies: In this paper, we show that phylogenetic divergence can be driven by different branching patterns that arise from specific ecological processes and propose a method to identify their signature in communities. Let’s picture two communities that experience different assembly processes (see schematic figure below), namely, competitive exclusion between close-relatives due to resource depletion (as predicted by limiting similarity theory, top community) and facilitation by a distant-relative nursery plant that mitigates the harshness of environmental conditions. Both mechanisms lead to increased phylogenetic divergence (more overdispersion), but the underlying branching pattern of such divergences (community phylogenies to the right) are markedly different. Still, the new communities may show similar phylogenetic divergence values, and therefore one may erroneously conclude that the same mechanism is at stake if the underlying branching patterns are ignored. Our method provides a handle to integrate both sources of information (i.e. phylogenetic divergence and the underlying branching patterns) using simple statistical tests.
Hypothetical plant communities experiencing different ecological processes, namely competitive exclusion between close-relatives (top) and facilitation (bottom). In the top-left community, resources are abundant and competition does not occur. When resources are scarce (top-right), competitive exclusion between close relatives comes into play and phylogenetic divergence increases. In the bottom-left, a community of species with narrow thermal niches thrive at an ambient temperature of 25ºC. A temperature increase of 5ºC (bottom-right) could lead to the collapse of the community, but the species can still persist under the canopy of a distantly-related facilitating species that provides microclimatic amelioration and augments phylogenetic divergence in the community.
Unexpected challenges: This research was not planned at all by the time I got involved in the Kili Research Unit, a multidisciplinary project that revolved around Mount Kilimanjaro biodiversity and ecosystem services. Dr. Markus Fischer, my postdoc advisor at the time, and Dr. Andreas Hemp, a botanist with more that 30 years of experience in the flora of East Africa, were interested in studying plant community assembly at Mount Kilimanjaro using phylogenetic information. However, I had a hunch that the tools available were insufficient for the project, so the opportunity presented itself allowing me to incorporate new ideas and concepts into existing theory. Through this I really had to put my statistics and programming skills to the test.
Major results and contribution to the field: Community phylogenetics is a young but controversial discipline, likely because too much has been demanded of both the original conceptual framework and classical descriptors of phylogenetic structure. In think our approach may contribute to mitigate this controversy by providing ecologists a handle to analyse phylogenetic divergence in the light of the underlying branching patterns, which is critical if we are to avoid spurious interpretations of phylogenetic information. I don’t mean by that our method is the ultimate solution, as the community phylogenetic discipline is not without methodological shortcomings that need addressing (see Cadotte et al. 2017, Ecological Monographs, 87, 535-551 for an excellent review), yet it represents one step forward in the field. To make the method more accessible to the community, we implemented it in R language and provided the code in full as a user-friendly function in the Supplementary of the article.
What are the next steps? There are still lots of interesting questions in community phylogenetics. For example, testing whether clades that are overrepresented in communities show different modes of trait evolution seems a promising avenue for future research (see Pearse et al 2019, Global Ecology and Biogeography, 28, 1499-1511 for a recent paper). Phylogenies are not magic wands that will unravel assembly mechanisms by means of few phylogenetic metrics, but just an important, exciting and necessary tool for understanding how biodiversity is generated and maintained. After all, elephants will never fly and butterflies will not eat lions because lineages are functionally constrained, meaning that evolutionary history matters. A new generation of eco-phylogenetic methods is coming up, and re-analyzing previous datasets with new available tools might unravel biological information that remains encoded in the phylogenies.
If you could study any organism on Earth, what would it be and why? I would love to delve into the flora of the Wallacea Islands (particularly Sulawesi and Moluccas) and New Guinea”. On the one hand, this region combines multiple biogeographically interesting factors such as tropicality, isolation (islands) and sharp environmental gradients (mountains), which make the region extraordinarily appealing to me. On the other hand, these islands awaken the sense of adventure that many biogeographers carry inside of us. Even today, there is a continuous dripping of new species of birds, small mammals and plants reported from New Guinea! I have already visited the region once, specifically the Wakatobi archipelago in southeast Sulawesi (mostly a diving trip, so that time was more about coral reefs, likely my favourite animal taxa), and I am determined to come back.
Any other little gems you would like to share? I love teaching. I have taught General Ecology and Botany in bachelor’s degree and phylogenetic methods in postgraduate courses so far, which complements my facet as a researcher. After all, today’s students will be tomorrow’s researchers, the reason why I consider teaching a fundamental duty of scientists. At this point, I have to make a confession; plants are my true motivation in science, and I could not imagine being an ecologist without a focus on plants. And guess what? This is simply because I had good botany teachers during my time as an undergraduate student.